

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Pre-eclampsia (PE) is clinical condition that occurs during pregnancy and is generally associated with an abnormal placenta [12]. PE is characterised by elevated blood pressure and proteinuria. It is one of the main causes of maternal and foetal mortality and affects women only after 20 weeks of gestation [345]. In developed countries, PE is responsible for approximately 15% of preterm deliveries and 18% of maternal deaths [678]. Despite extensive research, the underlying pathogenesis of pregnancy-induced hypertension (PIH) is not completely understood, and this limits the available treatment options [9].
Recent research has demonstrated that oxidative stress plays a critical role in the development of PE [10111213] and that increased levels of anti-angiogenic factors are significantly associated with PIH [141516]. This angiogenic imbalance is characterised by alterations in the ratios of pro-angiogenic factors, including vascular endothelial growth factor (VEGF) and anti-angiogenic factors, including fms-like tyrosine kinase-1 (Flt-1) and soluble VEGF receptor-1 (VEGFR-1). Remodelling of the maternal uterine vasculature occurs in patients with PE, and placental ischaemia and hypoxic conditions contribute significantly to angiogenic imbalance [1718]. The disrupted angiogenic balance and altered regulation of a variety of factors lead to inflammatory responses and oxidative stress, which eventually result in endothelial dysfunction and hypertension [192021]. Experimental studies using rodent models of PE have focused on restoring angiogenic balance, which can reduce hypertension [14152223]. As a result, identifying and developing therapeutic strategies that can reduce maternal inflammation, oxidative stress and endothelial dysfunction are extremely important for treating PE.
Pomegranate (Punica granatum) fruits are widely consumed and have antioxidant, anti-hyperglycaemic, hepatoprotective, cardioprotective and anti-epileptic effects [242526]. Pomegranate extracts can inhibit the proliferation of cancer cells [2728]. Pomegranate juice contains polyphenols including gallotannins, ellagitannins and flavonoids [29], which can improve cardiovascular activity, attenuate hypoxia-induced apoptosis, reduce inflammation and restore endothelial-cell function in vitro [3031]. Here we investigated the effects of punicalagin, the most abundant ellagitannin in pomegranate juice, on oxidative stress and angiogenic balance in rats with experimentally induced PE.
Go to : 
METHODS
Study animals
A total of 20 male (weight, 300–325 g) and 60 female (weight, 200–250 g) Sprague-Dawley rats were monitored in sterile cages under laboratory conditions (12 h day/night cycle; 22–23℃; humidity, 55–60%). The study was conducted in compliance with international guidelines for the study of laboratory animals and the national animal welfare laws of China. The experimental protocols were approved by the Southwest Medical University Animal Care Board [32].
Female rats in the same ovulation cycle stage were randomly separated into six sets (n=10 per group), where the set ratio was 3:1 females/males. The animals were bred for 3 days, and female rats were then inspected for the presence of a vaginal plug, which indicated successful copulation. The rats with vaginal plugs were separated and labelled ‘day 0 of pregnancy.’
Reagents and antibodies
Punicalagin and NG-nitro-L-arginine methyl ester (L-NAME) were obtained from Sigma-Aldrich (St. Louis, MO, USA). The buffers used for western blotting analysis were procured from the Beyotime Institute of Biotechnology (Beijing, China). The antibodies raised against VEGF and VEGFR-1/Flt-1 were from Cell Signaling Technology (Danvers, MA, USA), and the horseradish peroxidase (HRP)-labelled IgG secondary antibodies and β-actin were from Santa Cruz Biotechnology (Dallas, TX, USA). The remaining reagents used in this study were from Sigma-Aldrich unless otherwise stated.
Trial design
The animals were randomly separated into five groups of 12 rats. Hypertension was induced in pregnant rats using the nitric oxide synthase (NOS) inhibitor L-NAME [3334]. Each group of rats received 50 mg/kg/day of L-NAME by gavage on days 14–19 of pregnancy. The treatment group animals additionally received 25, 50 or 100 mg/kg of punicalagin every day by oral gavage on days 14–21 of pregnancy. During gestation, the pregnant rats were kept in an animal house under standard conditions.
On day 21 of gestation, the rats were anaesthetised using a mixture of intramuscular ketamine (90–100 mg/kg) and xylazine (10 mg/kg), and hysterotomies were performed. Both live and dead foetuses were collected and weighed. Placentas were also collected, and the rats were sacrificed by administration of intraperitoneal phenobarbital (100 mg/kg) after the surgery was completed.
Blood pressure measurements
Blood pressure was recorded on days 1, 7, 14, 16, 18 and 20 using the CODA 6 BP system (Kent Scientific, Torrington, CT, USA). The mean arterial pressure (MAP), systolic blood pressure (SBP) and diastolic blood pressure (DBP) were recorded.
Assessment of antioxidant capacity and oxidative stress
The placental tissue was homogenised in 0.1 M phosphate buffer and centrifuged. Supernatants were collected and analysed for antioxidants and oxidative stress markers. Protein concentrations were determined using the Bio-Rad bicinchoninic acid (BCA) assay kit (Bio-Rad, Hercules, CA, USA).
Antioxidant capacity: The antioxidant capacity of the placental tissue was measured using the Trolox equivalent antioxidant capacity (TEAC) assay kit (Cayman Chemical Company, Ann Arbor, MI, USA), in accordance with the manufacturer's instructions.
Measurement of thiobarbituric acid reactive substances (TBARS): TBARS are oxidative stress markers that bind to malondialdehyde. TBARS levels in placental tissue were determined using the TBARS assay kit (Cayman Chemical Company) in accordance with the manufacturer's instructions.
Protein carbonyl measurement: The levels of oxidised proteins in tissue can be used as an indicator of oxidative stress. Protein carbonyl levels in the placental lysates were measured using the protein carbonyl colorimetric assay kit (Cayman Chemical Company). Protein carbonyls react with 2,4-dinitrophenylhydrazine to form a Schiff base that produces hydrazone, which is measured using a spectrophotometer set to quantify absorption at 385 nm.
Catalase activity: The activity of the antioxidant enzyme catalase was measured in placental tissue using a commercial assay kit (Cayman Chemical Company) in accordance with the manufacturer's instructions. The data were recorded as U/mg of tissue protein.
Placental nitric oxide (NO)
Levels of NO in the peritoneal fluid were quantified using the nitrate/nitrite colorimetric assay kit (Cayman Chemical Company). This involves the conversion of nitrate to nitrite by nitrate reductase. The addition of Griess reagents then converts the nitrite to a deep-purple coloured azo compound, which is measured using an ELISA reader set to quantify absorption at 540 nm. The quantity of the azo compound accurately reflects the nitrite concentration of the original sample.
Reverse transcription polymerase chain reaction (RT-PCR) analysis
Total RNA was isolated from placental tissues and quantified using a BioPhotometer (Eppendorf, Hamburg, Germany) for RT-PCR analysis. First strand cDNA was synthesised using a cDNA reverse transcription kit (Applied Biosystems, Inc., Foster City, CA, USA). PCR was performed using a 7300 Real-Time PCR System (Applied Biosystems, Inc.) with SYBR green fluorescence. The following primers were used for amplification: VEGF-sense, 5′–ACCATGAACTTTCTGCTC–3′; VEGF-antisense, 5′–GGACGGCTTGAAGATATA–3′; VEGFR-1/Flt-1-sense, 5′–TTTGCATAGCTTCCAATAAAGTTG–3′; VEGFR-1/Flt-1-antisense, 5′–CATGACAGTCTAAAGTGGTGGAAC–3′; GAPDH-sense, 5′–CACCACCATGGAGAAGGC–3′, GAPDH-antisense, 5′–CCATCCACAGTCTTCTGA–3′. The PCR products were separated using 2% agarose gel electrophoresis. The bands were stained with 0.05% ethidium bromide and their intensities quantified using a Bio Gel imaging system (Bio Rad).
Immunoblotting
The placental tissues were homogenised on ice in radio-immune precipitation assay (RIPA) lysis buffer containing 50 mM Tris-HCl (pH 7.6), 1% NP-40, 0.5% sodium deoxycholate, 0.1% sodium dodecyl sulphate (SDS), phenylmethylsulphonyl fluoride (PMSF), 1 mg/L aprotinin and 1 mg/L leupeptin. The samples were centrifuged and the protein concentrations of the supernatants determined using the BCA assay kit (Bio-Rad). Equivalent quantities of protein (50 µg) from each group were separated using 10% SDS-polyacrylamide gel electrophoresis (PAGE). The separated protein bands were transferred to polyvinylidene difluoride (PVDF) membranes (Invitrogen, Carlsbad, CA, USA) and blocked for 60 min at 37℃ using Tris buffered saline with Tween-20 (TBST) buffer (20 mM Tris pH 7.6, 137 mM NaCl and 0.1% Tween 20) with 5% non-fat milk. The membranes were incubated overnight at 4℃ with specific primary antibodies (1:1,000) and then washed with TBST three times. Finally, the membranes were incubated at 37℃ for 60 min with HRP-labelled secondary antibodies (1:2,000). The positive bands were detected using an enhanced chemiluminescence detection kit (Merck Millipore, Burlington, MA, USA) and analysed using a ChemiDoc XRS imaging system (Bio-Rad). β-Actin was used as an internal control to normalise protein concentrations.
Statistical analysis
The experimental data were analysed using SPSS software (ver. 21.0; SPSS, Inc., Chicago, IL, USA). Group evaluations were performed using one-way analysis of variance (ANOVA) and Duncan's multiple range test (DMRT). A p-value<0.05 was considered statistically significant.
Go to :
RESULTS
Effect of punicalagin on blood pressure
Pregnant female rats that were treated with L-NAME had elevated SBP and DBP from day 14 through until day 20 (Figs. 1a–c). The mean SBP of L-NAME-treated rats was 172.6±2.32 mmHg on day 14 and 169.7±1.38 mmHg on day 20. These values were significantly higher than those recorded for the other experimental groups (p<0.05). The MAP measurements were similar to those recorded for SBP and DBP. The control group animals that were not treated with L-NAME had consistent BP measurements throughout the entire gestation period with only insignificant changes. The SBP and DBP measurements recorded for punicalagin-treated animals were significantly lower than those recorded for the L-NAME-alone group (p<0.05). Punicalagin-treated rats had significantly lower MAPs compared with the L-NAME-treated group from day 14 through until day 20. The highest dose of punicalagin (100 mg) decreased BP more effectively than did the lower doses of 25 and 50 mg.
 | Fig. 1Punicalagin decreased systolic (a) and diastolic (b) blood pressure and also decreased mean arterial pressure (c).Values are expressed as means±standard deviation (SD), n=6; one-way analysis of variance (ANOVA) followed by Duncan's multiple range test (DMRT) showed p<0.05; *p<0.05 vs. control; the letters a–e indicate mean values from experimental groups that differ significantly, p<0.05.
|
Punicalagin enhanced nitric oxide levels
The levels of vasodialator, NO in the placental tissues were measured. Placental NO concentrations were observed to be significantly (p<0.05) decreased in the L-NAME compared with the control-animal group (p<0.05; Fig. 2). Treatment with punicalagin at 25, 50 or 100 mg/kg resulted in dose-dependent increases in NO levels (p<0.05). 100 mg dose of punicalagin was found to be more effective compared to 25 or 50 mg doses. The NO levels observed in rats given the 100 mg-punicalagin dose were found to be near normal control group levels, indicating the efficacy of punicalagin.
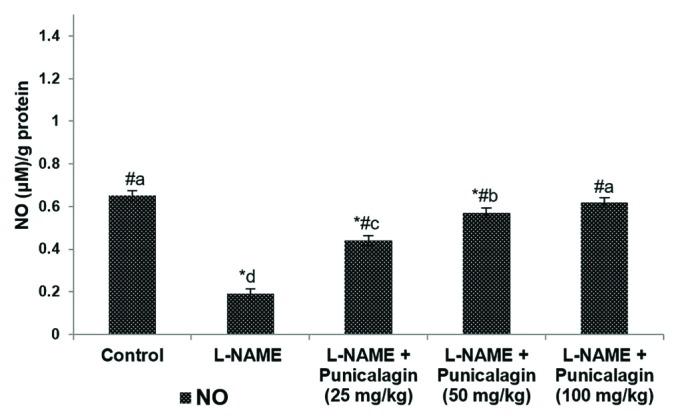 | Fig. 2Punicalagin increased placental nitric oxide (NO) levels.Values are expressed as means±SD, n=6; one-way ANOVA followed by DMRT showed p<0.05; *p<0.05 vs. control; #p<0.05 vs. NG-nitro-L-arginine methyl ester (L-NAME); the letters a–e indicate mean values from experimental groups that differ significantly, p<0.05.
|
Punicalagin reduced oxidative stress
Compared with the control group, the L-NAME-alone rats had a significantly decreased placental antioxidant capacity and significantly increased levels of TBARS and protein carbonyls (p<0.05; Figs. 3a–d). Additionally, the activity of the antioxidant enzyme catalase in the placental tissue had increased to a level comparable with the TBARS. Punicalagin administration considerably improved the TEAC, and decreased the levels of TBARS and protein carbonyls. Catalase activity decreased to almost normal levels following punicalagin treatment. These observations suggest that punicalagin reduced L-NAME-induced oxidative stress.
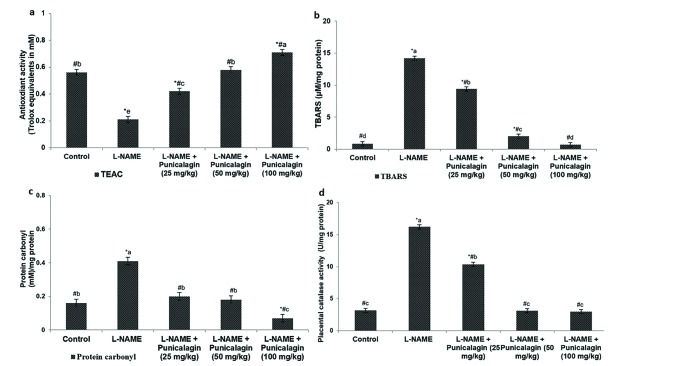 | Fig. 3Punicalagin enhanced the antioxidant capacity of the placenta (a), Punicalagin decreased the level of thiobarbituric acid reactive substances (TBARS) (b) and also decreased protein carbonyl levels in the placenta (c) and restored normal catalase activity levels (d).Values are expressed as means±SD, n=6; one-way ANOVA followed by DMRT showed p<0.05; *p<0.05 vs. control; #p<0.05 vs. L-NAME; the letters a–e indicate mean values from experimental groups that differ significantly, p<0.05.
|
Effects of punicalagin on PIH-related angiogenic imbalance
Angiogenic imbalance is strongly associated with the aetiology of hypertension and PE [35]. Administration of L-NAME to pregnant rats significantly decreased the expression of VEGF, both at the gene and protein levels (p<0.05). RT-PCR analysis revealed a marked decrease in VEGF mRNA levels in L-NAME-treated rats compared with the control group (Figs. 4a, b). The onset of PIH following L-NAME administration resulted in substantial increases in placental VEGFR-1/Flt-1 levels in the L-NAME-alone-treated rats compared with the control group.
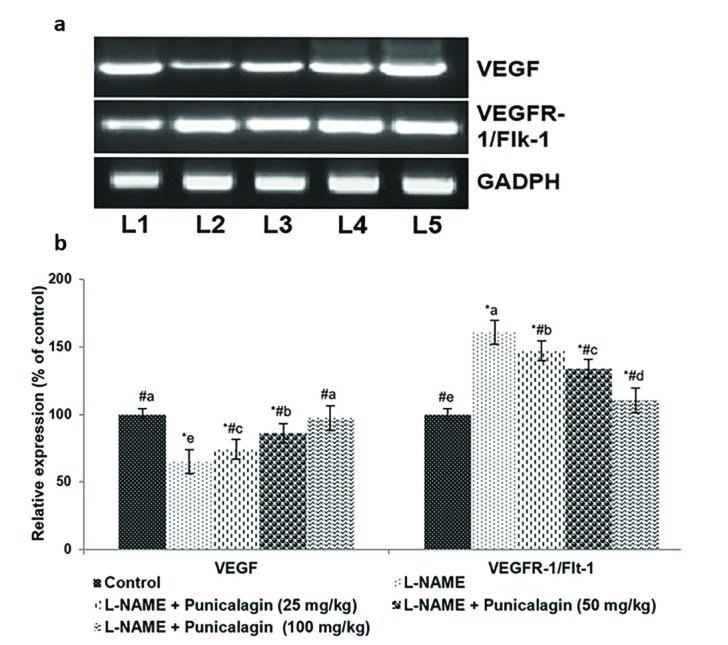 | Fig. 4mRNA levels (a) and protein expression (b) show that punicalagin restored angiogenic balance.Reverse transcription polymerase chain reaction (RT-PCR) analysis revealed a marked decrease in vascular endothelial growth factor (VEGF) mRNA levels in response to punicalagin treatment compared with control groups: L1, control; L2, L-NAME; L3, L-NAME+punicalagin (25 mg/kg); L4, L-NAME+punicalagin (50 mg/kg); L5, L-NAME+punicalagin (100 mg/kg). Values are expressed as means±SD, n=6; one-way ANOVA followed by DMRT showed p<0.05. Relative expression of proteins where the control expression level is set as 100%; *p<0.05 vs. control; #p<0.05 vs. L-NAME; the letters a–e indicate mean values from experimental groups that differ significantly, p<0.05.
|
Supplementation with 25, 50 or 100 mg/kg punicalagin caused a marked increase in VEGF mRNA levels and significantly decreased the levels of Flt-1 mRNA (p<0.05). The western blotting analyses also revealed significantly downregulated expression of VEGFR-1/Flt-1 and enhanced VEGF expression in response to treatment with punicalagin (Figs. 5a, b). These observations suggest that punicalagin effectively restored angiogenic balance.
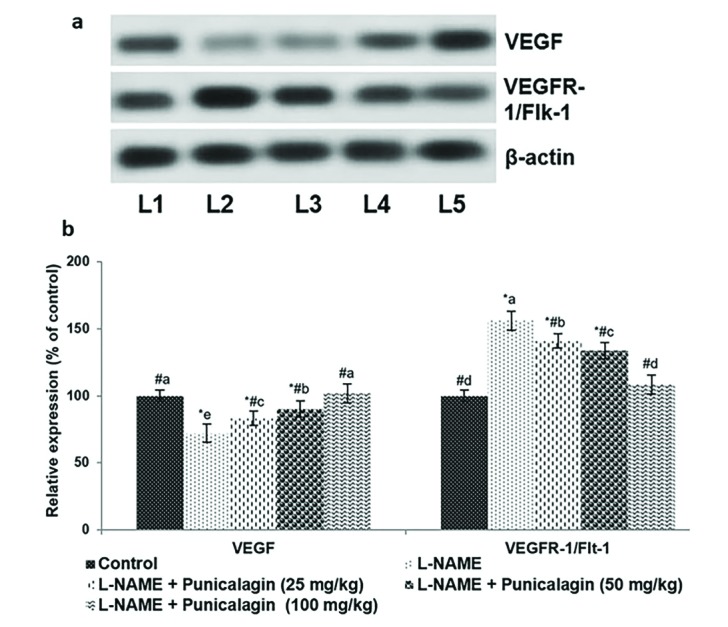 | Fig. 5Punicalagin restored angiogenic balance.Western blotting analysis (a) and protein expression (b) revealed a marked decrease in VEGF levels in response to punicalagin treatment compared with control groups: L1, control; L2, L-NAME; L3, L-NAME+punicalagin (25 mg/kg); L4, L-NAME+punicalagin (50 mg/kg); L5, L-NAME+punicalagin (100 mg/kg). Values are expressed as means±SD, n=6; one-way ANOVA followed by DMRT showed p<0.05. Relative expression of proteins where the control expression level is set as 100%; *p<0.05 vs. control; #p<0.05 vs. L-NAME; the letters a–e indicate mean values from experimental groups that differ significantly, p<0.05.
|
Go to :
DISCUSSION
PE is a hypertensive multisystem disorder and a principal cause of maternal and foetal mortality and morbidity [36] that affects 3–5% of pregnancies [37]. Although the pathophysiology of PE is not completely understood, the condition is generally associated with an abnormal placenta [12]. In this study, we investigated the effects of punicalagin in an L-NAME-induced animal model of PE. In humans, PE generally occurs during the third trimester, after 20 weeks of pregnancy, and our study was designed to reflect this. The gestation period of Sprague-Dawley rats is approximately 21 days; therefore, L-NAME was first administered on day 14.
L-NAME is an analogue of L-arginine. It competes at the active site of NOS, causing a reduction in NO synthesis that leads to higher blood pressure [38]. L-NAME can also induce PE-like symptoms, including proteinuria and growth retardation [3940]. In Sprague-Dawley rats, blood pressure higher than 129/91 mmHg is considered high. In this study, hypertension, the chief characteristic of PE, was induced using L-NAME and significant increases in SBP, DBP and MAP were observed. Following induction with L-NAME, blood pressure remained high throughout the gestation period. Additionally, placental NO levels decreased markedly following treatment with L-NAME.
NO generated by endothelial cells is reportedly involved in PE pathogenesis. NO plays an integral role in homeostatic vasodilation and contributes to vasodilatory responses during a normal pregnancy [41] In rats, the biosynthesis of NO is enhanced during pregnancy and stimulates vasodilation [42]. PE can decrease NO levels [4344]. We observed decreased levels of NO following treatment with L-NAME. The increases in SBP, DBP and MAP reflect the inhibitory action of L-NAME in reducing NO levels. Interestingly, the tested doses of punicalagin significantly enhanced NO levels and decreased blood pressure to almost normal levels. Previous studies with plant-derived compounds as resveratrol demonstrate anti-hypertensive effects and reduced oxidative stress [4546]. A triterpenoid, celastrol [47] and Epigallocatechin gallate (EGCG) from green tea [48] were found to enhance the effects of oral nifedipine, common anti-hypertensive drug in PE.
Oxidative stress plays a critical role in the pathophysiology of PE [49]. The data reveal an increase in oxidative stress in patients with PE, as well as in animal models of PE [505152535455]. Increases in the levels of free radicals produce oxidative damage in biomolecules, leading to cellular and endothelial dysfunction [56]. In this study, treatment with L-NAME resulted in elevated levels of placental malondialdehyde. The increased protein carbonyl content and decreased TEAC values reflected an increase in oxidative stress. Enhanced levels of placental catalase were also observed; this could be a protective measure to combat the excess of free radicals. Punicalagin decreased the level of placental oxidative stress, as reflected in the decreased levels of TBARS and protein carbonyls. This enhanced the total antioxidant capacity of the placenta and catalase activity was restored to levels similar to those measured in the control group.
Supplementation with antioxidants and essential fatty acids in pre-eclamptic women caused decreased oxidative stress and also aided in prevention or delayed onset of preeclampsia and as well improved the health of pregnant women [5758]. Studies in rodent model of PE showed that ω-3 PUFA supplementation reduced placental oxidative stress and enhanced fetal growth [59]. Studies demonstrated that supplementation with standard antioxidantsvitamin C and E markedly reduced oxidative stress and endothelial dysfunction [60] and pre-eclampsia [61]. The antioxidant efficacy of punicalagin could be responsible for reducing oxidative stress and improving TEAC.
Several studies have investigated the relationship between PE and altered placental vascular development [626364]. The signal protein VEGF is involved in both vasculogenesis and angiogenesis, which are critical for the development of the placenta [6566]. Angiogenic imbalance is frequently associated with PE [6768]. In patients with PE, the altered levels of VEGF and its receptors (VEGFR-1/Flt-1) disrupt angiogenesis, causing endothelial dysfunction and placental abnormalities [6970]. In our study, the significant decreases in VEGF expression and enhanced levels of Flt-1 that we observed following treatment with L-NAME showed that the angiogenic balance had been altered under hypertensive conditions. Downregulated expression of VEGF was observed at both the gene and protein levels. Punicalagin markedly upregulated VEGF expression and supressed VEGFR-1/Flt-1, restoring the angiogenic balance. Previous data demonstrated that resveratrol reduced soluble form of Flt-1 (sFlt-1) secretion from placenta that offers therapeutic potential in preeclampsia [71].
Several PE studies have reported decreases in blood pressure following restoration of angiogenic balance [1523]. Therefore, restoring angiogenic balance probably contributed to the decreases in blood pressure that we observed. These findings suggest that punicalagin can reduce hypertension by restoring angiogenic balance. Both the increases in NO levels and restoration of angiogenic balance could have contributed to the observed decreases in blood pressure.
Go to :
CONCLUSION
This study demonstrates that punicalagin can enhance NO levels, reduce oxidative stress and regulate the expression of VEGF and Flt-1, thereby reducing blood pressure in PIH. Punicalagin could potentially be used to treat PIH and PE. However, further studies will be required to comprehend the underlying molecular mechanisms involved.
Go to :
 XML Download
XML Download