

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Breast cancer accounts for approximately 23% of all cancers in Sri Lankan females and 12% of all cancers among Sri Lankans [1]. Notably, 62.1% of breast cancers in Sri Lankan women are diagnosed after the age of 50 years [1]. Such alarming figures highlight the importance of identifying individuals with higher risk of developing breast cancer, so that early detection, appropriate management, and preventive measures could be undertaken to reduce the morbidity and mortality rates associated with the disease [2].
Sporadic breast cancer is caused by a combination of environmental and genetic factors. Previous studies have shown that multiple genetic, epidemiological, and epigenetic factors contribute to its etiology [3]. For most sporadic breast cancers, a significant proportion of the risk is due to multiple low-penetrance susceptibility alleles [4]. In a recent study, common genetic polymorphisms in the XRCC2, PHB, CDH1, and ATM genes were found to be associated with increased risk of sporadic breast cancer among Sri Lankan postmenopausal women [5]. Several single nucleotide polymorphisms (SNPs) have been shown to be associated with clinicopathological profiles in sporadic breast cancer in major population groups [678910111213]. However, comparatively few studies have been conducted on under-represented South Asian groups, such as the Sri Lankan population [14].
Studies conducted on the Chinese population showed that the MDM2 SNP309 G allele increases the risk and the T allele is associated with earlier age of onset of sporadic breast cancers [10]. Similarly, a polymorphism in exon 11 of the BRCA1 gene, which leads to a glutamine-to-glycine substitution at codon 1,038, was found to be associated with early age of onset of sporadic breast cancer [13]. Chen et al. [15] observed that CD28 gene polymorphisms contribute to sporadic breast cancer risk and exhibit significant associations with clinicopathological features of sporadic breast cancer in the Chinese Han population. Xu et al. [12] reported that the rs11889031 CT genotype and the T allele in the inducible costimulator (ICOS) gene were associated with progesterone receptor (PR) status and lymph node metastasis in Chinese women.
Few descriptive studies have reported the clinicopathological features in Sri Lankan women with breast cancer. In a study conducted to document the breast cancer profiles of a group of Sri Lankan women, patient tumor characteristics and predicted prognosis were compared with immune profiles [16]. The overall profile of breast cancer and immune characteristics of Sri Lankan women were found to be largely comparable to profiles documented elsewhere in the South Asian region despite lower levels of expression of the estrogen receptor (ER). Similarly, in another Sri Lankan cohort, the expression levels of ER, PR, and human epidermal growth factor receptor 2 (HER2) were 53.2%, 50.0%, and 14.6%, respectively [17]. Lower tumor grade was significantly associated with ER and PR expression but not to HER2 amplification. Age at diagnosis was found to be significantly correlated with ER expression. In addition to these two descriptive studies, no published studies have comprehensively investigated the association between common genetic polymorphisms and the clinicopathological profiles in sporadic breast cancer in Sri Lankan women. Knowledge of such genetic variants will be beneficial for predicting breast cancer prognosis in the local population and will facilitate improved disease management.
The present study aimed to investigate the association between selected SNPs and clinicopathological profiles in sporadic breast cancer in Sri Lankan women. We conducted association testing for clinicopathological variables on a total of 350 women using single variant association testing strategies. A total of 58 SNPs in 36 breast cancer-related genes were investigated following a “case-control” strategy, with the categories defined by the presence or absence of the clinicopathological attribute. For example, patients whose tumors were HER2-positive were classified under one category (“cases”), while those who were HER2-negative were classified under another category (“controls”).
METHODS
Study design and study population
In the present study, ethylenediaminetetraacetic acid blood samples were collected between March 2010 and October 2011 from a Sri Lankan cohort comprising 350 unrelated postmenopausal women with histologically confirmed invasive breast cancer. Samples were obtained in accordance with the approval of the Ethics Review Committee of the Faculty of Medicine, University of Colombo (EC-15-082). Written informed consent was obtained from the study participants for future use of their samples for breast cancer-related genetic studies. Study participants were recruited postmastectomy and prior to administration of chemotherapy or radiotherapy. To exclude individuals with inherited cancer syndromes, any participant with a family history of any type of malignancy in their first and second degree relatives was excluded [5]. Clinicopathological patient data that were retrieved from an electronic database maintained at the Human Genetics Unit included the age of onset of cancer, tumor histologic grade, and lymph node involvement, as well as ER, PR, and HER2 status. ER, PR, and HER2 expression levels were evaluated following standard immunohistochemistry methods as part of routine diagnostic work. ER/PR expression was graded as follows: 0 (negative for receptor), 1+ (borderline), and 2+ to 3+ (positive for receptor). Samples were also scored based on the Allred scoring system: 0 (0% nuclear staining), 1+ (<10% nuclear staining), 2+ (10% to 75% nuclear staining), and 3+ (>75% nuclear staining). HER2 expression was graded as follows: 0 to 1+ (HER2-negative), 2+ (borderline/equivocal), and 3+ (HER2-positive). Samples were also scored based on the following scoring system: 0 (no staining is seen or membranestaining is <10% of the invasive cancer cells), 1+ (faint membrane staining in >10% of cells), 2+ (weak to moderate complete membrane staining in >10% of cells or <30% with strong, complete membrane-staining), and 3+ (strong, complete membrane staining in >30% of invasive cancer cells).
Selection of single nucleotide polymorphisms for genotyping
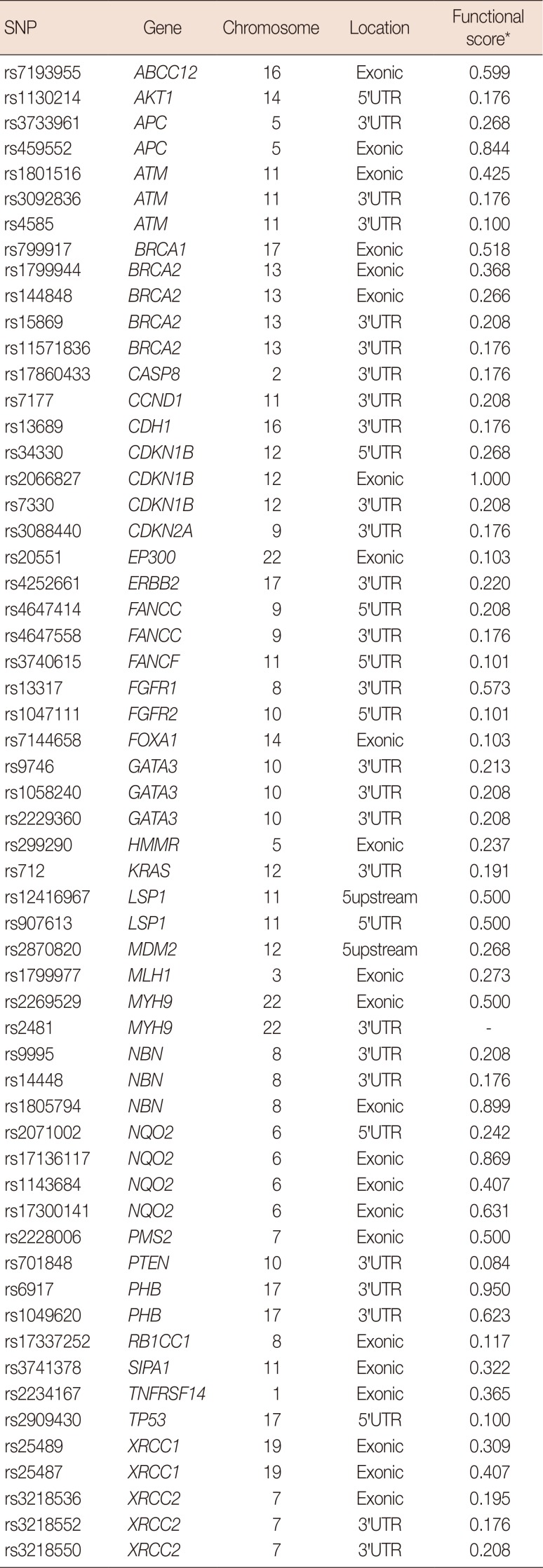
An extensive search of the published literature was carried out to identify the candidate genes involved in several cellular pathways that are involved in sporadic breast cancer. The SNPs in the candidate breast cancer genes that had minor allele frequencies greater than 0.05 in the HapMap Gujarati Indians in Houston, USA (GIH) were obtained from the International HapMap Project database. Genotype data from the Gujarati Indian population was used because they represent the only South Asian population group in the International HapMap project or other similar projects with dense genotypes available at the time of study design. The SNP nexus software tool (http://snp-nexus.org/) was used to select non-synonymous SNPs in the coding regions, 5′-untranslated regions (5′-UTRs), and 3′-untranslated regions (3′-UTRs) of the candidate genes. Coding synonymous and intronic SNPs were excluded [5]. The SNPs were further filtered using several bioinformatics software tools based on the validation status (dbSNP database; http://www.ncbi.nlm.nih.gov/SNP/), evolutionary conservation (MutationTaster; http://www.mutationtaster.org/), and presumed functional significance (SNP nexus; http://snp-nexus.org/) [18]. A total of 58 SNPs from 36 candidate genes that were associated with breast cancer were selected for genotyping (Table 1).
Genotyping and quality control
DNA extraction from all samples was performed using the Promega Wizard® Genomic DNA purification kit (Promega, Madison, USA) according to the manufacturers' protocol. The Promega QuantiFluor® dsDNA System (Promega) was used to quantify and normalize the samples to a concentration of 10.0 ng/µL. Genotyping of all the samples for the selected set of SNPs was performed using the Agena Bioscience Mass-Array technology on a Compact Spectrometer, iPLEX GOLD chemistry (Australian Genome Research Facility, Gehrmann Laboratories, University of Queensland, Brisbane, Australia). Primers flanking the gene regions containing the target SNPs were designed using MassArray Designer software [5]. A total of 57 SNPs was successfully genotyped with an average SNP call rate of 99.87%. SNP rs1047111 was excluded from analysis because the allele frequencies deviated from Hardy-Weinberg equilibrium at p-value <0.05. One sample was excluded, and a total of 349 cases passed filters and quality control.
Statistical analysis
Association testing was performed using logistic regression models adjusted for confounding factors using PLINK software version 1.07 (http://zzz.bwh.harvard.edu/plink/) [19]. The following covariates were included: body mass index, age at menarche, age at menopause, type of menopause (natural or artificial), age at first pregnancy, parity, breast feeding, use of hormonal contraceptives, use of hormone replacement therapy, and exposure to passive smoking (Table 2). Odds ratios (ORs) and their 95% confidence intervals (CIs) are presented along with p-values. Given that the present study is an exploratory study, p-value <0.05 was considered statistically significant.
RESULTS
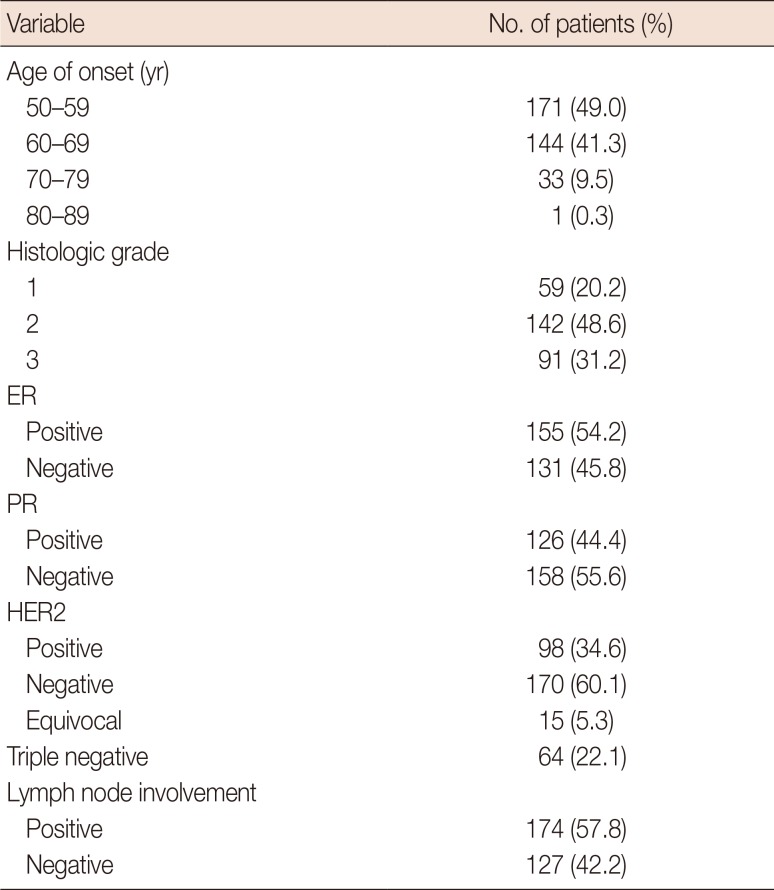
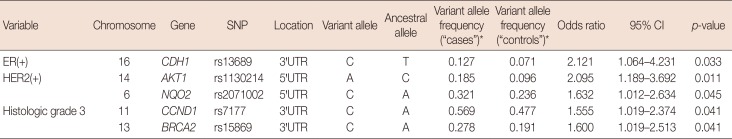
The mean and standard deviation of the age at onset of breast cancer was 60±7 years. The distribution of clinicopathological features in patients with breast cancer is shown in Table 3. The seven SNPs that showed significant associations with the clinicopathological profiles in breast cancer were as follows: rs799917 in BRCA1; rs17136117 in NQO2; rs13689 in CDH1; rs1130214 in AKT1; rs2071002 in NQO2; rs15869 in BRCA2; and rs7177 in CCND1. Detailed annotations of the SNPs that were significantly associated with breast cancer are shown in Table 4.
The G allele of BRCA1:rs799917 (p=0.047, β [standard error; SE]=−1.069 [0.537]) and the G allele of NQO2:rs17136117 (p=0.040, β [SE]=1.901 [0.923]) were associated with age of onset between 50 and 59 years. The C allele of CDH1:rs13689 (p=0.033; OR, 2.121; 95% CI, 1.064–4.231) was associated with ER-positive breast cancer. The A allele of AKT1:rs1130214 (p=0.011; OR, 2.095; 95% CI, 1.189–3.692) and the C allele of NQO2:rs2071002 (p=0.045; OR, 1.632; 95% CI, 1.012–2.634) were associated with HER2-positive breast cancer. The C allele of BRCA2:rs15869 (p=0.041; OR, 1.600; 95% CI, 1.019–2.513) and the C allele of CCND1:rs7177 (p=0.041; OR, 1.555; 95% CI, 1.019–2.374) were associated with high histologic grade (Table 5). No SNPs were found to be associated with PR-positive breast cancer, triple-negative breast cancer, and lymph node involvement in this cohort.
DISCUSSION
In the present study, we investigated the associations between SNPs located in the transcribed and regulatory regions of breast cancer-related genes with clinicopathological profiles in sporadic breast cancer in Sri Lankan women. Our study is the first to investigate the association between selected polymorphisms in breast cancer-related genes and the clinicopathological profiles in sporadic breast cancer in the Sri Lankan population. Among 56 tested SNPs, seven were found to be significantly associated with different clinicopathological profiles in sporadic breast cancer.
The G allele of rs799917 in exon 14 of the BRCA1 gene and the G allele of rs17136117 in exon 3 of the NQO2 gene were associated with age of onset between 50 and 59 years. BRCA1 is a key tumor suppressor gene that plays an important role in maintaining the stability and integrity of the genome. Previous studies have shown that the BRCA1 gene acts through various mechanisms, such as transcriptional repression, homologous recombination repair of double-strand breaks, and cell cycle control [20]. The C>T variation in rs799917 was reported to cause a proline-to-leucine change at position 871 in the BRCA1 protein, which constitutes a nonconservative change, given that proline confers unique structural properties to the polypeptide. PhastCons analysis of more than 28 species showed that the rs799917 variant is located in the middle of a highly conserved region of the BRCA1 gene. The BRCA1:rs799917 variant has been studied because it was found to be associated with sporadic breast cancer in several population-based studies [20]. However, most of the abovementioned studies reported conflicting and inconclusive results. Possible reasons include the nature of the study designs or differences in ethnicity and lifestyle factors, such as reproductive patterns, diet, and cultural practices [21]. Huo et al. [22] reported that rs799917 was associated with breast cancer risk in Chinese women. Nicoloso et al. [23] observed that the rs799917 T>C SNP, which is located in the BRCA1 coding region, affects the interaction between BRCA1 mRNA and miR-638. In turn, this leads to significant downregulation of BRCA1 expression among C allele carriers of rs799917, leading to increased risk of developing breast cancer. However, a meta analysis of 19,878 subjects by Qin et al. [3] reported that the BRCA1:rs799917 variant was not associated with breast cancer risk. Further studies are required to validate our findings.
On the other hand, excessive oxidative stress mediated by reactive oxygen species (ROS) can cause cellular deregulation, which in turn leads to cell proliferation, failure of cell apoptosis, and tumor promotion [24]. NQO2, an oxidative stress-related gene, protects cells from oxidative damage by catalyzing the reduction of carcinogenic quinone compounds into their hydroquinone forms. Furthermore, NQO2 is known to reduce the production of estrogen orthoquinones, as well as stabilize the p53 protein, a well-known breast tumor suppressor gene product [24]. However, no published studies have investigated the association between NQO2:rs17136117 and breast cancer. The variant NQO2 allele is likely to act by altered gene expression regulation mechanism, which in turn results in tumor promotion or downregulated expression or reduced activity of the ROS-reducing enzyme [25].
Steroid hormone receptors are valuable for the prediction and prognosis of breast cancer and are regarded as predictive markers for endocrine therapy [26]. The C allele of rs13689, which is located in the 3′UTR of CDH1 gene, was found to be associated with ER-positive breast cancer. The CDH1 gene, which encodes the calcium ion-dependent cell adhesion molecule E-cadherin, is known to play a crucial role in epithelial integrity and carcinogenesis [11]. In breast cancer, partial or total loss of E-cadherin expression leads to loss of differentiation characteristics, invasiveness, higher tumor grade, metastasis, and poor prognosis. Some studies have shown that the rs13689 variant is associated with breast cancer susceptibility [11]. In addition, the 3′UTR is important for mRNA stability and localization and potentially acts as the binding site of microRNA (miRNA). Polymorphisms in the 3′UTRs of several genes have been reported to be associated with diseases because of their effects on miRNA-regulated gene/protein expression [11]. The rs13689:C allele is likely to exert an effect through one of these mechanisms but no published studies have reported on the association between this variant and ER-positive breast cancer.
The C allele of rs2071002 and the A allele of rs1130214 in the 5′UTR of the NQO2 and AKT1 genes, respectively, were found to be associated with HER2-positive breast cancer. The ancestral A allele of rs2071002 abolishes the binding site of transcriptional-activator Sp1 [25]. On the other hand, the C allele is known to create a new Sp1 binding site, which is associated with increased activity of the NQO2 gene promoter in normal human breast cells [25], thereby causing stronger expression of NQO2 in normal breast and peri-tumoral tissues relative to wild-type tissues. No published studies have investigated the association between the variant allele and HER2-positive breast cancer.
AKT1 is a serine-threonine kinase that is regulated by the phosphatidylinositol 3-kinase (PI3K) signaling pathway that is known to influence apoptosis, cell proliferation, migration, and survival [27]. Atypical activation of the PI3K-AKT-mTOR (PAM) signaling pathway has been reported in hormone-related tumors, such as breast and ovarian cancers [27]. Activated AKT may initiate a series of biological events involving survival, adhesion, motility, and proliferation, thus promoting malignant transformation of cells and tumor progression. The A allele of rs1130214 has been postulated to act by causing aberrant expression of genes involved in the AKT signaling pathway. This has potential clinical implications because abnormal activation of the PAM pathway is known to be closely correlated with the efficacy of medical treatment for breast carcinoma, including endocrine therapy, anti-HER2 therapy, and chemotherapy [27]. In early or advanced HER2-positive breast cancer, PAM pathway activation has been reported to be associated with recurrence, metastasis, poor prognosis, and low overall survival. Notably, a recent preclinical study demonstrated that inhibition of the PAM pathway promoted antiproliferative activity in various breast cancer cell lines, including HER2 overexpressing cells that are resistant to trastuzumab and lapatinib [28]. Thus, AKT1 gene polymorphisms could serve as prognostic and therapeutic biomarkers for breast cancer and warrant further investigation and validation.
The C allele of rs15869 and the C allele of rs7177 in the 3′UTR of the BRCA2 and CCND1 genes, respectively, were found to be associated with high histologic grade (Grade 3). The 3′-UTR polymorphisms that disrupt miRNA binding are well-known to exert functional roles and act as genetic markers for cancer risk [29]. SNPs located at the miRNA-binding sites of double strand break repair genes, such as BRCA2, can influence cancer risk by dysregulating the expression of target genes. Studies have demonstrated that rs15869, which is located at a miRNA binding site in the BRCA2 gene, is associated with higher breast cancer susceptibility. A Chinese study reported that rs15869 together with age at menarche and parity could increase the risk of breast cancer by 2.39-fold [30]. However, the association between rs15869 and tumor grade has not previously been reported and thus requires further investigation.
CCND1 is known to play a major role in cell cycle control, especially in the transition from the G1 to the S phase, which is regulated by cyclin-dependent kinases [9]. Mutations and overexpression of CCND1 is known to lead to alterations in cell cycle progression and is frequently observed in a variety of tumors and could thus contribute to tumor progression. However, pooled analysis of case-control studies from the Breast Cancer Association Consortium showed that the rs7177:C allele is associated with reduced breast cancer risk in European women [9]. The association between rs7177 and high tumor grade in breast cancer has not previously been reported in other populations.
In conclusion, the findings of this exploratory study suggested that SNPs located in the AKT1, BRCA1, BRCA2, CCND1, CDH1, and NQO2 genes are associated with different clinicopathological profiles in sporadic breast cancer among Sri Lankan women. The lack of association of the remaining SNPs could be explained by the small effect sizes or the relatively small sample size of the present study. Another possibility could be that these SNPs are low penetrant alleles that exert their effects through complex gene-gene and/or gene-environmental interactions. Additionally, the power of detection may have been limited by missing data in some of the clinicopathological variables that were evaluated. Our findings have potential implications in guiding health professionals for improved prediction of the prognosis and effectiveness of treatments for sporadic breast cancer. However, further studies are required to replicate these findings in other population groups and delineate the exact biological mechanisms underlying the effects of the target SNPs.


 XML Download
XML Download