

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Atopic dermatitis (AD) is a type of skin inflammation that is caused by both genetic and environmental factors1.
Human mannose-binding lectin (MBL) is a serum lectin taking part in the innate immunity by binding various microorganisms and activating the lectin-complement23. Opsonization defect due to deficiency of MBL is associated with increased susceptibility to infection and stems from the presence of a low efficiency promoter and/or three gene mutations in exon 1 (variants B, C, and D) of MBL223. The patients with abnormal homozygous MBL alleles are susceptible to an immune-deficiency disease that is not related to HIV4.
MBL deficiency could trigger AD or complications when they are additionally exposed to an infection. The first evidence of MBL involvement in AD was suggested by a family study which showed that children with low plasma MBL levels who were homozygous for allele B of MBL were prone to pruritic skin disease and possibly AD, with or without recurring infections5. In addition, a report revealed that low MBL levels were clearly associated with the BB MBL2 haplotype in a Turkish family members who also had recurrent skin infections and AD5. Moreover, the three exon 1 variants B, C, and D of MBL were more frequently observed in Brazilian AD patients than in healthy controls, although the disease severity was not investigated6. In this study, children with AD had higher frequency of allele O of MBL gene related to low or deficient levels of MBL compared to the control group6. Likewise, low or deficient serum MBL levels may result in predisposition to AD7, although there was a conflicting report that showed no association of the B allele of MBL with AD susceptibility in a Japanese population8.
We hypothesized that the modulation of innate immune defense by MBL2 variants influenced a wide range of susceptibility to AD. Therefore, we investigated whether single-nucleotide polymorphisms (SNPs) of the MBL2 gene could be proper genetic diagnostic factors in Korean AD patients by examining the SNPs and haplotypes, including −550 and −221 in the promoter region, +4 in the 5′ UTR and codon 54 in exon 1 of the MBL2 gene.
MATERIALS AND METHODS
Subjects
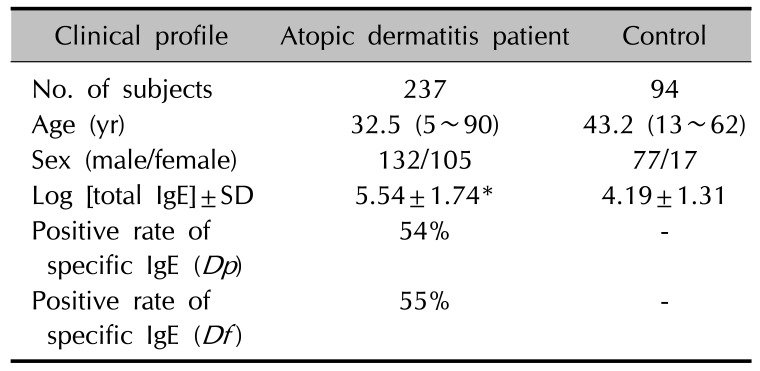
In this study, we included 237 unrelated Korean AD patients (132 males and 105 females; mean age 32.5±18.0 years; age range 5~90 years) who were registered in the Department of Dermatology, Uijeongbu St. Mary's Hospital in Korea. All patients showed moderate to severe AD according to the Hanifin's criteria9. Controls were 94 healthy persons without a personal or family history of AD. For genetic studies, blood was collected by venipuncture, and genomic DNA was prepared using QIAamp blood kit (QIAGEN, Hilden, Germany). Blood [total immunoglobulin E, IgE] was measured by LPIA-200 system (Iatron Corp., Tokyo, Japan). Blood IgE levels were in the range of 2~50,000 IU/ml (median [25th percentiles~75th percentiles], 282.5 [87.1~954.0]). Assays for specific IgE antibodies to Dermatophagoides pteronyssinus (Dp) and Dermatophagoides farina (Df) were performed with the Pharmacia CAP FEIA immunoassay on a UniCAP 100 automatic analyzer (Pharmacia and Upjohn, Uppsala, Sweden) according to the manufacturer's directions. An antigen-specific IgE value of over 0.35 kU/L was classified as increased. The clinical data are presented in Table 1. This study was carried out from March 3, 2003 to December 25, 2004 in compliance with the principles of the Declaration of Helsinki. Since there was no statutory law during that time, only verbal consent was obtained from the patients and healthy persons after explaining the purpose of our study and their rights. In case of children were included in the study, the parent or guardian was informed orally and they agreed to the purpose and procedure of our study. The coded research data obtained from March 3, 2003 to December 25, 2004 were reanalyzed and the Institutional Review Board (IRB) of Uijeongbu St. Mary's Hospital approved this study on November 12, 2015 (IRB no. UC15RISI0160).
Molecular methods
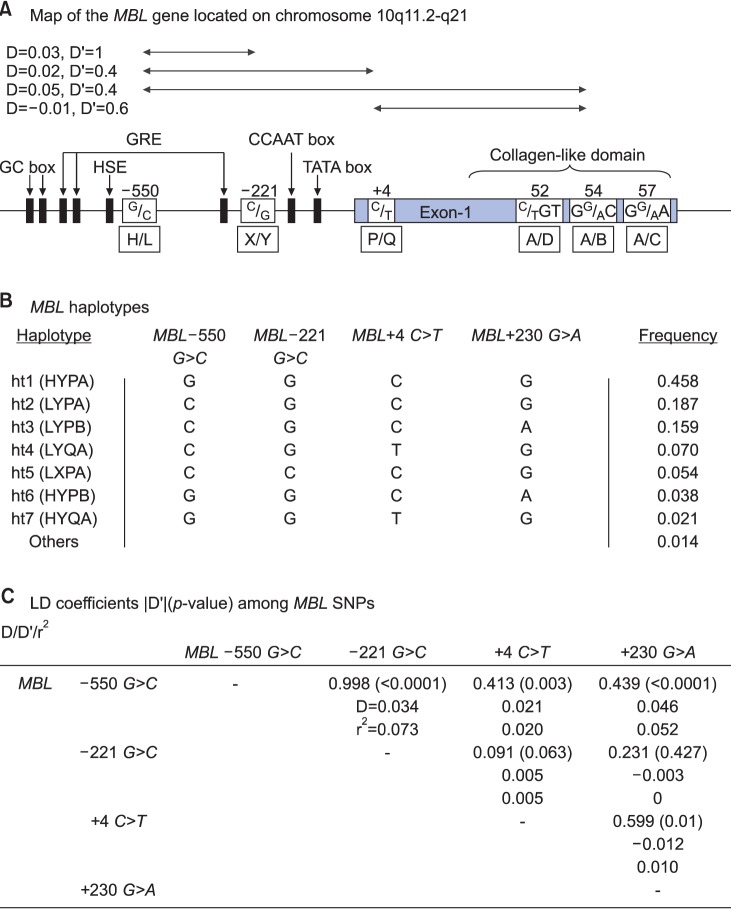
Primer sets were designed for the four polymorphic sites: H/L at −550, X/Y at −221, P/Q at +4, and A/B at codon 54 (Fig. 1A), and used for differential PCR analysis. Genotyping of the promoter region at position −550 (rs11003125) of the MBL2 gene and codon 54 (rs1800450) of exon 1 was carried out by the polymerase chain reaction (PCR)-restriction fragment length polymorphism (RFLP) analysis as described before1011. Genotyping of the promoter region at position −221 (rs7096206) of the MBL2 gene was performed by the PCR-sequence specific primer method, as described by Steffensen et al.12. DNAs with one point mutation in the 5′ UTR at position +4 (P/Q variants) of the MBL2 gene were amplified by site-directed mutagenesis (SDM)-PCR13. The P and Q alleles were detected by RFLP performed on the SDM-PCR products using a mutated 5′-primer (Table 2)10111213. AccI cleaved the 261 bp PCR product specific for the L allele into two fragments (239 bp and 22 bp), while BanI cleaved the A and B alleles. SacI and HindIII cleaved the 136 bp PCR products specific for the P and Q alleles into 110 bp and 26 bp fragments, respectively. PCR restriction fragments were separated by 8% polyacrylamide gel electrophoresis.
Statistics
Hardy-Weinberg equilibrium was analyzed for the gene frequencies obtained by simple gene counting and the chi-square test was performed for comparing the observed and expected values. We examined a widely used measure of linkage disequilibrium (LD) between all pairs of bi-allelic loci, Lewontin's D′ (|D′|)15. Haplotype frequencies were calculated using the Phase 2.0 program, as described elsewhere16. Phase probabilities of each site were also calculated for each individual by the Phase 2.0 program. Individuals with phase probabilities less than 0.987 were excluded from the analysis. Genetic effects of inferred haplotypes were analyzed in the same way as SNPs. Comparison of genotype and haplotype frequencies for each MBL2 polymorphism was carried out by chi-square test. Odds ratios (ORs) with 95% confidence intervals (CIs) were calculated using SAS ver. 8.1 (SAS Institute, Cary, NC, USA). p<0.05 after Bonferroni's adjustment for multiple testing of the four SNPs in the diplotype was considered to be statistically significant throughout the study. An OR provides an effect estimate, whereas a score of <1 is related to a protective effect, and a score of >1 is related to an increased risk. Genotype distribution of MBL2 SNPs and haplotypes among AD patients and normal controls were analyzed with logistic regression models adjusted for age, sex and log [total blood IgE] levels as covariates.
RESULTS
Characteristics of the study population
The clinical characteristics of 331 subjects are shown in Table 1. Mean age was higher in the controls compared to the patients, and males were predominated in both groups. Around 54% of the patients with AD were positive for Dp-specific IgE and 55% of patients were positive for Df-specific IgE. Subjects with AD had higher blood log [total IgE] levels (Student's t-test, p<0.0001).
Allele frequencies of the MBL polymorphisms
The MBL2 gene located on chromosome 10q11.2-q21 consists of four exons with a total size of ~10.0 kb. Four SNPs in the MBL2 gene (Fig. 1A); three within the upstream of the promoter region (a G-C transversion at position 550, a C-G transversion at position 221, and a C-T transition at position +4 in the 5′ UTR) and one G-A transition at +230 calculated on the codon 54 of exon 1 were genotyped by the single-base extension method. As shown in Fig. 1, MBL −550 G>C, −221 G>C, +4 C>T and +230 G>A were included in the statistical analysis. All four SNPs were in complete (|D′|=1 and r2≠1) or absolute LD (|D′|=1 and r2=1). Seven haplotypes were identified without any ambiguous phasing due to LDs among SNPs (Fig. 1B, C).
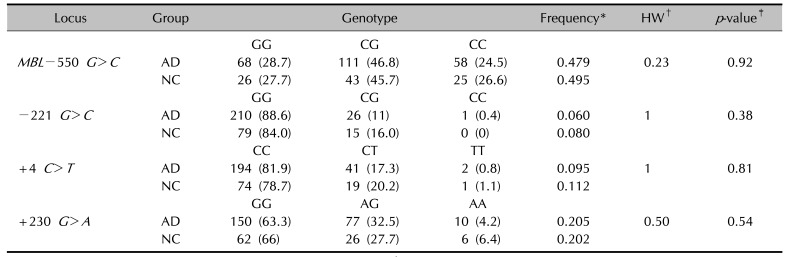
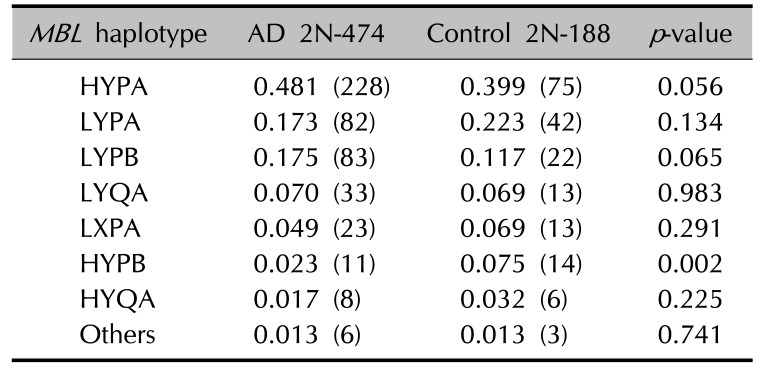
Allele distributions of the polymorphisms on MBL2 (MBL −550 G>C, −221 G>C, +4 C>T, and +230 G>A) were determined in the study population (Table 3). No significant difference was found in the MBL2 polymorphism frequencies between AD patients and controls.
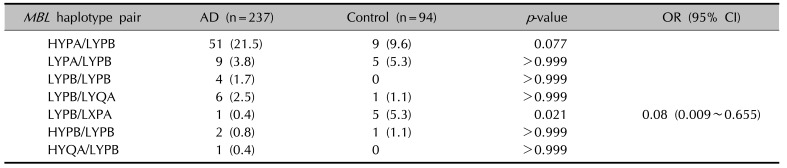
When haplotypes with a frequency greater than 0.01 were analyzed among the AD patients and controls, 7 haplotypes were observed at positions −550 and −221 in the promoter region, at +4 in the 5′ UTR, and at codon 54 in exon 1 of the MBL2. The frequency of the MBL2 HYPB haplotype was significantly decreased (p=0.002) in the AD patients (Table 4). The frequencies of the heterozygous genotype MBL2 LYPB/LXPA (OR, 0.08; 95% CI, 0.009~0.655; p=0.021) were also significantly decreased in the AD patients (Table 5).
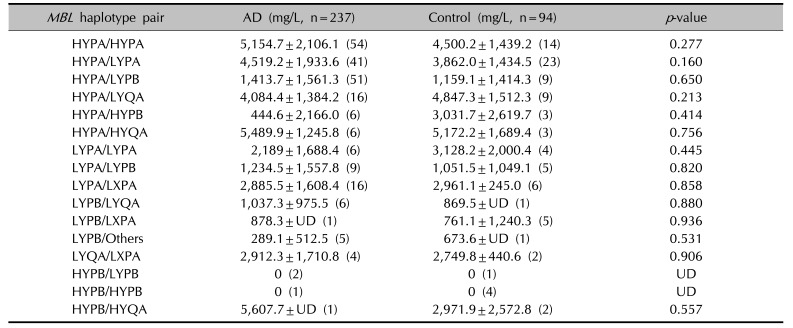
The blood MBL levels were not significantly different between the AD patients and controls (mean±standard deviation: 3.30±2.40 vs. 3.06±2.05, respectively, p=0.077, Student's t-test). Interestingly, the blood MBL level was not detected in both AD patients and controls with the MBL2 HYPB/HYPB and HYPB/LYPB haplotypes, possibly leading to deficiency of MBL (Table 6).
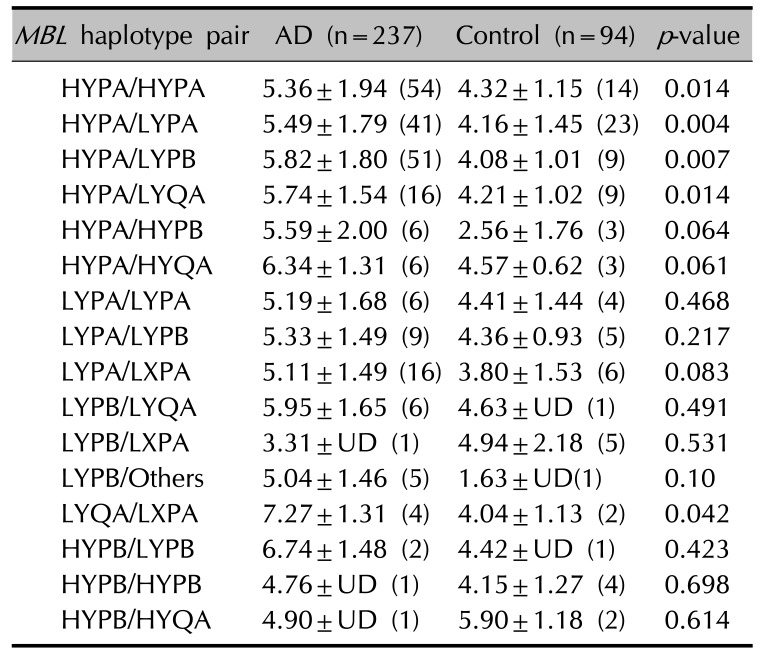
The blood MBL levels were significantly correlated with total IgE levels in the AD patients (r=0.13, p=0.048, Pearson's correlation). In addition, the blood log [total IgE] of MBL2 HYPA/HYPA, HYPA/LYPA, HYPA/LYPB, HYPA/LYQA, and LYQA/LXPA haplotype pairs were significantly increased in the AD patients compared to controls (Table 7).
DISCUSSION
In this study, we found a significant decrease in the frequency of MBL2 haplotype HYPB, −550G (H), −221G (Y), +4C (P), and codon 54Asp (B) in Korean AD patients. We also found that blood log [total IgE] levels of MBL2 HYPA/HYPA, HYPA/LYPA, HYPA/LYPB, HYPA/LYQA, LYQA/LXPA haplotype pairs were significantly increased in the AD patients.
In a previous study, higher frequency of point mutation at codon 54 (B) of the MBL gene has been implicated as a candidate susceptibility loci in Brazilian AD patients 7. We found that the frequency of MBL HYPB/HYPB haplotype was decreased in the AD patients, although the plasma MBL was not detected in both AD patients and controls with this haplotype and the number of subjects was too small for the statistical analysis (Table 6). MBL deficiency would not be a susceptibility factor for the development of certain complications. For example, it was demonstrated that AD patients with Eczema herpeticum (EH), a systemic complication caused by herpes simplex virus infection had similar levels and functional activities of serum MBL as AD patients with no history of EH17. Considering the sample size limitations in this study and our study, further research is needed to determine the detailed defects that predispose subjects to viral infection and AD.
Previous studies have demonstrated that there are large differences in the frequencies of exon 1 variant and haplotypes of MBL2 in various ethnic groups. For example, the codon 54 (B) mutant was found with a higher frequency in in Caucasians (0.22~0.28) and in Asians including Japanese and Korean patients (0.11~0.26), but very rare in Africans (0.0~0.03)11121819. In addition, the HYPA haplotype, the most effective MBL-producing haplotype, was found in Asians including Koreans (0.44~0.47) and Caucasians (0.25~0.34) and commonly in Eskimos (0.81). In contrast, the LXPA haplotype was found rarely in Asians including Koreans (0.07~0.10) and Eskimos (0.03), but it was found more commonly in Caucasians (0.18~0.26)3111218.
Various genetic polymorphisms in the MBL gene have been reported as risk factors for the development of a certain clinical subtype and severity. The susceptibility to Behcet's disease (BD) is related to a higher frequency of the MBL2 HYPA haplotype, which may cause increased acute or chronic hyper-inflammatory responses and influence the severity of BD19. A significantly higher percentage of patients with BD showed high serum MBL levels (≥500 ng/ml) compared to controls and were associated with skin lesions in Korean patients20. In addition, the progression of systemic lupus erythematosus is associated with MBL gene polymorphism and serum MBL concentration21.
In our study, the blood MBL levels were significantly correlated with total IgE levels in the AD patients. In addition, blood log [total IgE] levels of MBL2 HYPA/HYPA, HYPA/LYPA, HYPA/LYPB, HYPA/LYQA, and LYQA/LXPA haplotype pairs were significantly increased in the AD patients. Extrinsic AD shows high total plasma IgE levels and has specific IgE for environmental and food allergens, whereas intrinsic AD shows normal total IgE levels and the absence of specific IgE22. Our previous study demonstrated that the polymorphisms of the macrophage migration inhibitory factor (MIF) promoter related to innate immunity were significantly associated with an increased risk for AD. Especially, the −794 7-CATT locus and the MIF C/7-CATT haplotype were significantly associated with decrease of total IgE levels in the blood, suggesting that these polymorphisms might be a marker for intrinsic AD23. Therefore, the HYPA haplotype pair of the MBL2 gene, which is related to higher total blood IgE levels, would be a possible marker for extrinsic AD.
In conclusion, we investigated the SNPs and the haplotypes of the MBL2 gene, which might be proper genetic diagnostic factors in Korean AD patients. The frequency of MBL2 LYPB/LXPA had a possibly protective effect in Korean AD patients.
 XML Download
XML Download