

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Keloids are characterized by hyperproliferative growth of dermal fibroblasts and excessive collagen deposition in the dermis12. Previous studies suggested that the initial step in the development of the fibrotic reaction in keloids involves the expression of transforming growth factor β1 (TGF-β1) in neovascular endothelial cells, thus inducing the adjacent fibroblasts to express markedly high levels of TGF-β1/β2 and their receptors, as well as type I and VI collagen34. Recent studies emphasize a potential role for TGF-β1 intracellular signaling pathways, especially for the Smad pathway, in the pathogenesis of keloids4. Smad2 and Smad3 are overexpressed and the inhibitory Smad6 and Smad7 are reduced in keloid fibroblasts135.
Treatments for keloids include surgical excision, intralesional corticosteroids, 5-fluorouracil, bleomycin and interferon, topical imiquimod, compression, cryotherapy, radiation, silicon sheeting and laser or light-based therapies, showing variable success. Recurrence is common, even with combination therapy6. Therefore, preventing keloid is an important issue.
Low-level light therapy (LLLT) uses low levels of visible to near infrared light. It has been widely used to reduce pain and inflammation, to promote wound healing, and to prevent tissue necrosis. Many reports demonstrate that LLLT modulates normal dermal fibroblasts and the TGF-β family1. Recently, two clinical reports showed that LLLT was effective in preventing hypertrophic scars and keloids78. However, studies investigating the effect of LLLT on keloid fibroblasts are still rare, especially the ones investigating the effects of LLLT at molecular level. Herein, we aimed to evaluate the effect of LLLT using blue (410 nm), red (630 nm), and infrared (830 nm) light on the collagen synthesis in keloid fibroblasts, by observing the signaling molecules involved.
MATERIALS AND METHODS
Isolation and culture of keloid fibroblasts
Keloid fibroblasts were isolated from keloid-revision surgery samples after obtaining informed consents from the patients. Keloid fibroblasts were cultured in Dulbecco's modified Eagle's medium (DMEM), supplemented with 10% fetal bovine serum, 100 U/ml penicillin and 100 µg/ml streptomycin at 37℃ in a humidified atmosphere containing 5% carbon dioxide (CO2).
Irradiation
Keloid fibroblasts were seeded in a 6-well plate at 3.0×105 cells/well. APL® (Medro Medical Division, Seoul, Korea) was used for this study. This device produces blue (410 nm), red (630 nm), and infrared (830 nm) light emitting diode (LED) light. The power of each wavelength was 205 mW/cm2 for blue, 172 mW/cm2 for red, 50 mW/cm2 for infrared. Keloid fibroblasts were irradiated twice, with the second irradiation 24 hours after the first one. Preliminary experiment was performed to determine the lowest fluence to inhibit the proliferation of keloid fibroblasts. Cells were maintained in 400 µl Dulbecco's phosphate-buffered saline during irradiation and transferred in DMEM after irradiation for 24 or 48 hours.
Cell viability assay
To determine the direct effect of LLLT on the proliferation of viable cells, we counted the number of viable cells using the EZ-CyTox cell viability assay kit (Daeillab, Seoul, Korea). Twenty-four hours after the last irradiation, the cells were treated with 40 µl of Ez-CyTox solution and incubated for additional 3 hours at 37℃. Absorbance was measured at 450 nm using an ELISA reader (Molecular Device, Menlo Park, CA, USA). Viability was expressed as percentage of viable cells compared to the non-irradiated control. The experiment was repeated twice with two set of fibroblasts.
Real-time quantitative reverse transcription polymerase chain reaction
The mRNA of collagen type I, TGF-β1, Smad3, and Smad7 were measured in 24 and 48 hours after last irradiation. Total RNA was isolated using RNAasy® mini kit (Quiagen, Hilden, Germany). cDNA transcription was performed using the Superscript® III First-Strand (Invitrogen, Carlsbad, CA, USA) according to the manufacturer's instructions. cDNA was analyzed by real-time quantitative reverse transcription polymerase chain reaction using ABI Prism 7000 Sequence Detection System (Applied Biosystems, Foster, CA, USA). Fluorescent signals were collected during extension phase, Ct values of the sample were calculated, and the transcript levels were analyzed by 2−ΔΔCt method. Primers and internal probes for collagen type I, TGF-β1, Smad3, and Smad7 were purchased as assays on demand primer-probe sets (Applied Biosystems). The experiment was repeated five times.
Western blot analysis
The protein of collagen type I, TGF-β1, Smad3, and Smad7 were measured in 48 hours after last irradiation. Cells were lysed in lysis buffer (50 mM Tris [pH 7.4], 150 mM NaCl, 0.1% NP 40, 1% sodium dodecyl sulfate [SDS], 0.5% deoxycholic acid, 1 mM EDTA, protease and phosphatase inhibitors). Samples were separated on 10% SDS-polyacrylamide gels and transferred onto polyvinyldenefluoride membranes. The protein blots were incubated with antibodies against collagen type I (Abcam, Cambridge, MA, USA), TGF-β1 (GeneTex, Irvine, CA, USA), Smad3 (Epitomics, Burlingame, CA, USA), Smad7 (Santa Cruz Biotechnology, Santa Cruz, CA, USA) and actin (Santa Cruz Biotechnology). Detection was performed via enhanced chemiluminescence (Amersham Biosciences, Buckinghamshire, UK). The intensity of each band was quantified using Image J and the data were reported as relative intensity according to actin.
Statistical analysis
IBM SPSS Statistics Desktop ver. 20.0.0 (IBM Co., Armonk, NY, USA) was used for statistical analysis. Results are expressed as mean and standard deviation. Mann-Whitney U test was used for statistical evaluation between control and experimental groups in the study. A p-value lower than 0.05 was considered statistically significant.
RESULTS
Determination of the fluence of LLLT
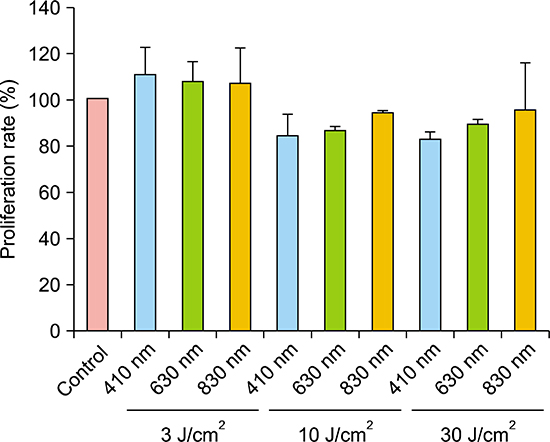
Keloid fibroblasts were irradiated once with 3, 10, 30 J/cm2 fluences of the three wavelengths of 410-, 630-, and 830-nm as preliminary experiment. Tryptan blue assay exhibited proliferations rates of keloid fibroblasts tend to decrease only at 10 and 30 J/cm2, showing rather increased proliferation rate at 3 J/cm2, in all wavelengths, although it was not significant (Appendix). In the condition of similar proliferation rate, using lower dose is more suitable and convenient for conducting experiment. Therefore, we determined to irradiate 10 J/cm2 for keloid fibroblasts.
Cell viability assay
Viable keloid fibroblasts decreased with irradiation with all wavelengths compared to non-irradiated control. However, it was not statistically significant (Fig. 1).
mRNA expression of TGF-β/Smad signaling components
Collagen type I expression was significantly decreased with irradiation at 410 nm in 48 hours (p<0.005). We also observed lower Smad3 expression after irradiation at 410 nm in 48 hours, but it was not statistically significant (p=0.065). There were no differences between irradiated samples in 24 hours and non-irradiated controls (Fig. 2, Table 1).
Protein levels of TGF-β/Smad signaling components
Irradiation at 410 nm reduced the protein levels of collagen type I in 48 hours. However, we observed no differences between irradiated samples and the control for TGF-β1, Smad3, and Smad7 (Fig. 3).
DISCUSSION
In the present study, we investigated whether LLLT could affect the collagen synthesis in keloid fibroblasts. Irradiation with 410 nm decreased collagen type I expression in keloid fibroblasts.
In normal fibroblasts, the effect of LLLT can be explained by photobiomodulation. Wavelengths above 500 nm are widely used to stimulate wound healing9. Their energy is absorbed by the cytochrome C oxidase in the mitochondrial membrane and activates various cellular signaling pathways through inducing the production of nitrogen oxide, adenosine triphosphate and reactive oxygen species (ROS)10. It can induce the transcription of various growth factors including platelet-derived growth factor, TGF-β, and fibroblast growth factor, and cytokines such as interleukins and tumor necrosis factor-alpha. On the other hand, blue light induces a higher production of intracellular ROS in a dose-dependent manner, which might act as a mediator of cellular effects of blue light11. Because of their relative close wavelengths, it is possible that blue and ultraviolet A light has similar biological effect such as ROS generation and immunomodulation11. Flavins are photo-acceptors and involved in the generation of ROS. Flavins are small, water-soluble molecules that absorb wavelengths below 500 nm and initiate free radical reactions when excited by light. Flavin-mediated photosensitization generates cytotoxic free radicals and is responsible for significant biological effects such as mutagenesis1213.
Previous reports indicate that 410 nm LED irradiation is cytotoxic for normal fibroblasts. Opländer et al.14 demonstrated that single irradiation with 410 or 420 nm LED at 5~10 J/cm2 inhibits the proliferation of dermal fibroblasts. Bonatti et al.15 also reported a significant reduction in the number of normal fibroblasts irradiated with 470 nm light at 18 J/cm2. According to Seo et al.16, red (630 nm, 9.5 J/cm2) and green (530 nm, 9.8 J/cm2) LED-irradiated cells were significantly more proliferated than cells irradiated with blue light (460 nm, 27 J/cm2). Also, blue light did not increase the expression of collagen type I and TGF-β1 in the dermis compared to the control, whereas red and green light enhance the expression of both genes.
In contrast to studies on normal fibroblasts, reports on the effect of LLLT on keloid fibroblasts are extremely rare and disappointing. Bonatti et al.15 showed that a single irradiation with 470 nm LED at 6, 12, and 18 J/cm2 does not induce significant differences in the number of keloid fibroblasts. To our knowledge, no study reports that LLLT affects the molecules investigated here in keloid fibroblasts.
Our results showed that collagen mRNA levels decreased after irradiation in keloid fibroblasts, which means that LLLT inhibited the transcription of this gene. Unfortunately, it is uncertain whether the TGF-β/Smad signaling pathway is inhibited in keloid fibroblasts, because other downstream signaling components, except collagen type I, did not show any significant changes in their expression after LLLT. It should be considered that LLLT also affects other signaling pathways in keloid, such as the mitogen-activated protein kinase (MAPK), which is suggested to mediate TGF-β/Smad signal transduction1718. The influence of MAPK on keloid fibroblasts was evaluated after pulsed dye laser (PDL) treatment in a few reports. Activation of extracellular signal-regulated kinase and the p38 signal transduction pathway is involved in the suppression of keloid fibroblast proliferation and the induction of apoptosis after PDL treatment1920. Recently, possible role of wingless type (Wnt)/β-catenin signaling pathway in keloid pathogenesis has been reported, which was known to play a key role in cellular proliferation. Wnt5a and β-catenin, its effector, were highly expressed in keloid fibroblasts and mediate the effect of TGF-β in wound healing process21. In addition, unlike normal fibroblasts, abnormal TGF-β signaling and autocrine loop are important in keloid fibroblasts14. For example, keloid fibroblasts show marked sensitivity to TGF-β, causing an accelerated increase in collagen and fibronectin synthesis compared to normal fibroblast2223. Therefore, the peak time of the TGF-β/Smad signaling components in keloid fibroblasts might be different from that of normal fibroblasts and it might be difficult to predict their expression pattern14.
This study has significance, in that it demonstrated that LLLT with 410 nm light inhibited collagen synthesis in keloid fibroblasts in vitro. Considering the initial pathophysiology of keloid, in which the expression of the TGF-β1 activates the adjacent fibroblasts to produce collagen, LLLT might be effective in preventing keloid formation at the initial stage. Corresponding clinical studies with blue LED and further investigative studies are needed to demonstrate the signaling mechanisms in keloid fibroblasts after LLLT. We hope this study can be a starting point for clinical application of LLLT to patients with keloid scars.



 XML Download
XML Download