

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Eosinophils actively participate in the pathogenesis of allergic skin diseases.12 Extensive extracellular deposition of eosinophil major basic protein has been observed in the lesional skin of atopic dermatitis and chronic urticaria.34 A correlation has been demonstrated between eosinophil numbers, eosinophil cationic protein concentrations, or both, and the severity of atopic dermatitis.567 Eosinophil granule proteins possess cytotoxic activities,8 suggesting a potential mode of eosinophil contribution to dermal tissue damage in allergic skin diseases. Nevertheless, mechanisms responsible for the dermal infiltration of eosinophils remain unclear.
Immunoglobulin E (IgE) has been the most convincing candidate for the induction of eosinophilic skin inflammation because serum IgE levels are elevated in >80% of atopic dermatitis patients. Approximately 85% of patients have positive immediate skin test or radioallergosorbent test (RAST) results for specific IgE antibodies (Abs) in response to various allergens.910 Mast cells bearing IgE for the relevant antigen release eosinophil-activating mediators.11 Involvement of IgE in tissue eosinophilia was demonstrated using animal models of asthma.12 Nevertheless, concentrations of IgE are raised in most but not all patients with atopic dermatitis and do not necessarily correlate with disease severity.1314 Animal studies demonstrating the non-essential role of humoral immunity in the development of eosinophilic inflammation also contradict the role of IgE.15161718 Taken together, the importance of IgE in the pathogenesis of eosinophilic inflammation in atopic dermatitis has become controversial.
Activated T-helper type (Th) 2 cells and Th2 cytokines are crucial for the infiltration and activation of eosinophils.1920 The number of CD4+ T cells was increased in the skin lesions and peripheral blood of atopic dermatitis patients.21 In atopic dermatitis, the infiltrating CD4+ T cells appear to have a Th2 phenotype.22 A Th2 cytokine, interleukin (IL)-5, drives terminal differentiation of committed eosinophil precursors while also promoting eosinophil activation, chemotaxis, and prolonged survival,23 suggesting that IL-5 plays a central role in the induction of eosinophilic inflammation. However, it is not certain whether Th2 cells have the potential to induce atopic dermatitis-like eosinophilic skin inflammation without the assistance of IgE-mediated immune responses.
This study was performed to delineate the role of Th2 cells in the development of allergic eosinophilic inflammation in the skin by employing T-cell transfers in mice.
Go to : 
MATERIALS AND METHODS
Animals and in vitro polarization of T cells
Mice expressing DO11.10 TCRαβ, which recognizes residues 323-339 of ovalbumin (OVA), are described elsewhere.24 Male BALB/c mice at 6 to 8 weeks of age were obtained from Japan SLC, Inc. (Hamamatsu, Japan). OVA-specific Th1 and Th2 cells were generated from DO11.10 mice, using methods described previously.2526 Successful differentiation of polarized cells was achieved using enzyme-linked immunosorbent assay (ELISA) and the intracellular staining of produced cytokines.2526 Experimental procedures used in this study were approved by the Animal Use and Care Committee of Tokyo Metropolitan Institute of Medical Science.
Cell transfer and challenge procedures
The dorsal skin of BALB/c recipients was shaved with electric clippers and then depilatory cream was applied (Kanebo, Tokyo, Japan). Twenty-four hours later, the mice were anesthetized with inhaled halothane (1% v/v), and a dorsal shaved skin area 6 mm in diameter was irradiated (900 µW/cm2 at 254 nm) with an ultraviolet lamp (TK-151; Unicom, Chiba, Japan) for 1 minute. OVA-specific Th1, Th2, or naive DO11.10 T cells were stained with the fluorescein-based dye 5 (and 6)-carboxyfluorescein diacetate succinimidyl ester (CFSE; Molecular Probes, Eugene, OR, USA) by methods described previously.2627 Then, 3×107 cells were injected into the tail vein of wildtype BALB/c mice. Twenty-four hours after the transfer, an aluminum rivet 9 mm in diameter was bonded to the shaved/irradiated area with cyanoacrylate adhesive (Toa Gousei, Tokyo, Japan) before being gently peeled off 10 minutes later. Then, a polypropylene cup-skin chamber 9 mm in diameter containing a sterile cotton pellet (30 mg) with 0.3 mL OVA solution (300 µg/mL in heparinized saline, 10 U/mL) was bonded to the area. Control mice were challenged with bovine serum albumin (BSA). For inhibition studies, dexamethasone (Dex) and FK506 (FK) were suspended in saline containing 0.5% Tween-80 (Tokyo Kasei, Tokyo, Japan) and administered subcutaneously 30 minutes before OVA challenge.
Skin-chamber fluid (SCF)
Twenty-four hours after antigen challenge, mice were killed by overdose with ether anesthesia before the skin chamber was removed from the skin. The cotton pellet was disentangled using 2 mL of Hank's balanced salt solution (HBSS) containing 0.1% BSA for 1 hour at room temperature. The SCF was then transferred to a test tube and centrifuged (250×g) for 5 minutes at 4℃.
Cytokines in the resulting supernatant were determined by ELISA, employing rat anti-mouse IL-5 monoclonal Ab (mAb) (BD Biosciences, Franklin Lakes, NJ, USA) and anti-mouse MCP-3 Ab (R&D systems, Minneapolis, MN, USA) as the capture Abs and biotinylated rat anti-mouse IL-5 mAb and goat anti-mouse MCP-3 Ab (R&D systems) as the detecting Abs. IL-4 and interferon (IFN)-γ were assayed by Duo Set ELISA Development Systems (R&D systems), MCP-1 by OptEIA mouse MCP-1 ELISA set (BD Biosciences), eotaxin by Quantikine mouse Eotaxin ELISA kit (R&D systems), and regulated on activation, normal T cell expressed and secreted (RANTES) by AN'ALYZA mouse RANTES immunoassay kit (R&D systems), each according to the manufacturer's instructions. The minimum detectable concentrations were 5, 5, 20, 5, 2, 5, and 50 pg/mL for IL-4, IL-5, IFN-γ, eotaxin, RANTES, MCP-1, and MCP-3, respectively.
The remaining cell pellet was immediately suspended in 250 µL of HBSS and the total cell number in the SCF was counted using an automatic cell counter (pocH-100iV; Sysmex, Hyogo, Japan). The number of CFSE-positive cells was counted using a FACSCantoII flow cytometer (BD Bioscience). Additional differential cell counting was performed by using microscopy on centrifuged preparations stained with May-Giemsa, counting 200 cells per animal.
Tissue eosinophil peroxidase (EPO) activity and histology
EPO activity in the skin was measured as previously described28 but with slight modifications. Briefly, after removing the skin chamber, the underlying skin was isolated by punch biopsy (10 mm diameter), and cut into fragments with a scalpel. Tissue fragments were suspended in 2 mL of 0.05 M Tris buffer (pH 8.0) containing 0.5% hexadecyltrimethylammonium bromide (Tris-HTAB) and homogenized using a Polytron homogenizer (Kinematica, Luzern, Switzerland) for 30 seconds. Homogenates were centrifuged (2,000×g) for 20 minutes at 20℃ then the supernatants were passed through a 0.45-µm filter. One volume of substrate solution (0.04% H2O2, 1.2 mg/mL o-phenylenediamine dihydrochloride in Tris-HTAB) was added to the filtered homogenate and mixed for 5 minutes at 20℃. The reaction was terminated by adding the same volume of 4 M H2SO4 and the absorbance at 492 nm was measured. Tissue EPO activity was expressed in units using standard horseradish peroxidase, whose specific activity was known.
The skin tissue was fixed in 10% neutral buffered formalin and paraffin-embedded. Sections (4 µm) were stained with hematoxylin and eosin for histologic analysis.
Statistical analysis
Data are presented as mean±standard error. Statistical analysis was performed by using Student's t test and one-way analysis of variance with Bonferroni's method. A P value of <0.05 was considered to indicate statistical significance.
Go to :
RESULTS AND DISCUSSION
Whether Th2 cells are capable of recruiting eosinophils into the skin without the assistance of antigen-specific IgE was first investigated. CFSE-labeled Th1/Th2-differentiated or naive DO11.10 T cells were infused into unprimed mice, followed by OVA challenge. Twenty-four hours later, the number of CFSE+ cells (antigen-specific T cells) and eosinophils in the SCF as well as EPO activity in the skin were measured. In Th2 cell-transferred mice, the number of both eosinophils and antigen-specific T cells in the SCF was increased upon challenge with OVA, in comparison with that in BSA (Fig. 1A). In Th1 cell-transferred mice, antigen-specific T cells migrated into the SCF to an extent similar to that in Th2 cell-transferred mice, while antigen-induced eosinophil accumulation was much weaker. Infiltration of neither T cells nor eosinophils occurred in the mice transferred with naive T cells. In parallel with eosinophil accumulation, EPO activity in the skin was specifically induced in Th2 cell-transferred and OVA-challenged mice, suggesting that EPO activity is a reliable indicator of eosinophil recruitment into the tissues (Fig. 1A).
 | Fig. 1induced migration of T cells and eosinophils in the skin tissues of OVA-reactive T cell-transferred mice. Naive or Th1/Th2-differentiated DO11.10 T cells (3×107) were stained with CFSE and transferred to wildtype mice by intravenous injection. After 24 hours, these mice were challenged with 300 µg/mL OVA or BSA. Twenty-four hours after challenge, the CFSE-positive, antigen-specific T cells and eosinophils in SCF and EPO activity in the skin tissues were measured (A). Data are expressed as the mean±SEM (n=5-6). Four days after challenge, a skin specimen was taken, stained with hematoxylin-eosin, and observed under optical microscopy (B). The low and high magnification images shown in the upper and lower panels, respectively, are representative of three-four animals. Representative eosinophils are indicated by arrow heads. Bar=50 µm. OVA, ovalbumin; Th, T-helper type; CFSE, 5 (and 6)-carboxyfluorescein diacetate succinimidyl ester; BSA, bovine serum albumin; SCF, skin-chamber fluid; EPO, eosinophil peroxidase; SEM, standard error of mean. *P<0.05; †P<0.01 (Student's t test).
|
Antigen-induced skin inflammation was evaluated in histological sections. In naive T cell-transferred mice, inflammatory cells were detectable only in small numbers even upon challenge with OVA (Fig. 1B). Consistent with the results of cell infiltration and tissue EPO activity, various inflammatory cells, including lymphocytes and neutrophils, but not eosinophils, migrated into the skin of Th1 cell-transferred mice (Fig. 1B). Obvious inflammatory features similar to the symptoms of atopic dermatitis patients, including massive accumulation of eosinophils, were seen in the skin of Th2 cell-transferred mice upon OVA challenge.
In these short-term experimental conditions, OVA-specific IgE was not detected in the skin or sera (data not shown), suggesting that eosinophilic skin inflammation could be induced by Th2 cells without assistance of IgE. Although hyper-IgE is a common feature of patients with atopic dermatitis,910 a dispensable role of IgE in the development of allergic inflammation was also confirmed in other target tissues, such as lungs, nasal mucosa, and intestine, in which massive eosinophil accumulation was seen in Th2 cell-transferred mice.252930 Although obvious eosinophilic accumulation was not observed in OVA-specific IgE-transgenic mice even upon repeated antigen challenge,30 we have shown that the IgE/mast cell-dependent pathway plays a supplemental role in eosinophilic inflammation, by employing an antigen-immunized mouse model.17 The cooperation of Th2 cells and IgE in the pathology of atopic dermatitis deserves further investigation.
To elucidate mechanisms by which eosinophils and T cells migrate into the skin, cytokine concentrations in the SCF of T cell-transferred mice were next examined (Table). In accordance with the in vitro profiles of transferred T cells,2526 IL-4, and IL-5 were specifically produced upon antigen challenge in the mice transferred with Th2 cells, whereas IFN-γ was preferentially produced in Th1 cell-transferred mice. Eotaxin concentration in the SCF production was increased only in Th2-transferred mice upon antigen challenge, whereas RANTES was produced only in Th1-transferred mice. MCP-1 and MCP-3 levels were increased by antigen challenge in both Th1 and Th2 cell-transferred mice.
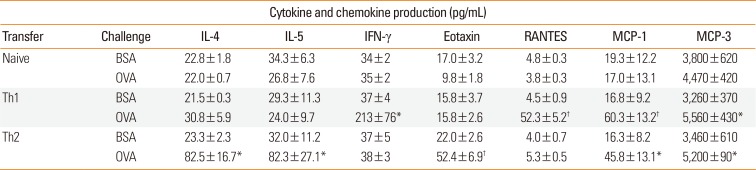
Table
Antigen-induced cytokine and chemokine production in the SCF of DO11.10 T cell-transferred mice
Naive or Th1/Th2-differentiated DO11.10 T cells (3×107) were transferred to wildtype mice by intravenous injection. After 24 hours, these mice were challenged with 300 µg/mL OVA or BSA using a skin chamber. Twenty-four hours after challenge, the concentrations of cytokines and chemokines in the SCF were measured.
Data are expressed as the mean±SEM (n=5-6).
SCF, skin-chamber fluid; IL, interleukin; IFN, interferon; RANTES, regulated on activation, normal T cell expressed and secreted; MCP, monocyte chemoattractant protein; BSA, bovine serum albumin; OVA, ovalbumin; Th, T-helper type; SEM, standard error of mean.
*P<0.05; †P<0.01, compared with BSA-challenged control (Student's t test).
![]()
Eosinophils express high levels of CC chemokine receptor (CCR) 3,3132 which binds several chemokines, including eotaxin, RANTES, and MCP-3. These chemokines are released from various tissues and cells in response to cytokine stimulation.33 It is intriguing that a significant recruitment of eosinophils was not seen in the skin of Th1 cell-transferred mice, in which substantial production of RANTES and MCP-3, as well as migration of Th1 cells themselves, were observed. A Th2-specific cytokine, IL-5, promotes eosinophil chemotactic activity.3435 The cooperation of IL-5 and eotaxin in the accumulation of eosinophils has been demonstrated in vivo.36 Therefore, IL-5 activity might be important for the eosinophilic skin inflammation selectively induced by Th2 cells. Further studies are needed to elucidate the mechanisms and meaning of our new finding that eosinophil-active RANTES and MCP-3 are produced in Th1-mediated pathology without recruiting eosinophils.
To gain an insight into the relationship between the antigen-induced infiltration of eosinophils and T cells that occurred in Th2 cell-transferred mice, the effects of the immunosuppressants Dex and FK were next examined. As shown in Fig. 2, both agents similarly and dose-dependently suppressed antigen-induced infiltration of antigen-specific T cells and elevation of skin EPO activity in Th2 cell-transferred mice. These results support the clinical efficiency of Dex and FK for atopic dermatitis patients. Since these reagents strongly suppress activation of T cells but not eosinophils,373839 it is suggested that skin-infiltrating, antigen-specific Th2 cells participate in the accompanying eosinophil migration.
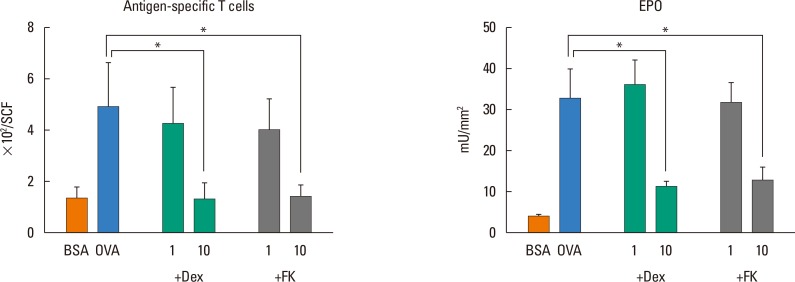 | Fig. 2Effects of Dex and FK on antigen-induced Th2 cell migration and upregulation of EPO activity in the skin. Th2 cells (3×107) were transferred to wildtype mice by intravenous injection. After 24 hours, these mice were challenged with 300 µg/mL OVA or BSA. Indicated doses of Dex and FK were administered subcutaneously 30 minutes before challenge. Twenty-four hours after challenge, the number of CFSE-positive cells in the SCF and EPO activity in the skin tissues were measured. Data are expressed as the mean±SEM (n=5-6). Dex, dexamethasone; FK, FK506; Th, T-helper type; EPO, eosinophil peroxidase; OVA, ovalbumin; BSA, bovine serum albumin; CFSE, carboxyfluorescein diacetate succinimidyl ester; SCF, skin-chamber fluid; SEM, standard error of mean. *P<0.05 (Bonferroni's method).
|
In conclusion, Th2 cells have the potential to develop eosinophilic skin inflammation, a representative pathological feature of atopic dermatitis, independent of an IgE-dependent pathway. Our new model of skin infiltration of antigen-specific Th2 cells and eosinophils in Th2 cell-transferred mice responds to Dex and FK and is expected to be useful for the generation of novel therapeutic treatments for atopic dermatitis, especially targeting Th2 cell-mediated and eosinophil-related pathogenesis.
Go to :
 XML Download
XML Download