

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Atopic dermatitis (AD) is a chronically relapsing inflammatory skin disease. Impairment in the skin barrier function plays an important role in the development of AD, along with an altered cutaneous immune-response associated with recurrent skin infection and allergen sensitization.12
It has been widely accepted that genetic susceptibility is closely associated with the occurrence and prognosis of AD.3 With regard to the impairment in skin barrier function, variants including in the filaggrin gene (FLG), serine protease inhibitor (SPI) Kazal-type 5 gene (SPINK5), and serine protease (SP) Kallikrein 7 gene (KLK7) play key roles in AD occurrence.456 The prevalence of FLG mutations in the Caucasian population is relatively high, present in 40% of Northwestern European AD patients.7 AD patients with FLG mutations show earlier onset age, greater disease severity, and a higher tendency to progress to the atopic march.8 However, in contrast to the prevalence in Caucasian AD subjects, recent studies have shown that the frequency of FLG mutations is relatively lower in the Asian population.9 Meanwhile, genetic polymorphisms in SP and SPI have also been found to be closely related to the onset of AD.10 With the SPINK5 variation, the flawed inhibition of SP results in hyperactivity of SP, which leads to impairment in skin barrier function.11 However, in terms of SPI and the SPINK5 mutation, only scarce data has been published, especially in an Asian population.12 According to our previous report, some variations including single nucleotide polymorphisms (SNPs) in KLK7, SPINK5 and FLG are associated with the occurrence of AD.13
Due to the deficiency in antimicrobial peptides (AMPs) represented by defensin, AD patients are more susceptible to bacterial, fungal, and viral skin infections than the normal population. In addition to β-defensin 1 (DEFb1), kinase insert domain receptor (KDR) and tumor necrosis factor-α (TNF-α) are well known innate immune response-related genes in AD.1415 Human DEFb1 plays a crucial role in suppressing bacterial and candidal colonization,16 and TNF-α is an important mediator between innate and adaptive immunity.1417
KDR is associated with allergen sensitization in AD.14 According to our previous report, heterozygous type variations (Hetero) in DEFb1 (rs574339) and KDR (rs2305948) are related with AD occurrence.13
Along with innate immunity, adaptive immunity also plays an important role in the development of AD.18192021222324 According to recent studies, Fc epsilon receptor 1 alpha (FcεR1α) and interleukin (IL)-4, 5, 9, 10, 12, 13, and 18 are particularly important in the development of AD. The elevated expression of FcεR1α facilitates antigen presentation to dermal dendritic cells and enhances T cell priming.25 It is well known that the Th2 dominant immune response plays an important role in AD. Immunoglobulin E (IgE) is a marker of allergic sensitization, and IL-4 and 13 play a pivotal role in the IgE-dependent inflammatory reaction. In addition, IL-3, 5, 9, and 10 act on mast cell, basophil, and eosinophil growth and maturation, and IgE, IL-12, and 18 have a well-known correlation with AD.26 Numerous studies have reported that a variety of IL-associated inflammatory responses are associated with AD.1819212223 Our previous report showed Hetero in IL-5RA (rs334809), IL-9 (rs31563), IL-12RB1,2 (rs393548, rs436857), and IL-13 (rs20541) are associated with AD occurrence.13
Treatments of AD are classified into basic, active, proactive and adjunctive treatments.27 In general, AD that shows favorable response to basic and topical treatments can be expected to have a good prognosis. If not, however, active treatment with systemic drugs should be considered. In particular, immunesuppressants such as cyclosporine, methotrexate, or azathioprine may need to be used for an extended period of time in those with treatment-resistant diseases, which has a significant mental and economic burden on both the patient and family and can adversely affect the quality of life.
The clinical severity of AD is mostly based on the eczema score observed at a single time point. However, in a more longitudinal view, even severe patients at a single time point can be divided into whether or not their disease can be easily controlled with basic management including antihistamines and topical corticosteroids, and various immunomodulators such as methotrexate or cyclosporine are not needed for treatment. If genetic factors are associated with the treatment response of AD, it would be useful to establish a future treatment plan and to enhance the patient's understanding of the treatment process before starting AD therapy. Based on these hypotheses, we performed this study to predict the recalcitrant disease by clarifying the clinical characteristics and genetic variations in AD subjects. By predicting whether AD patients will become recalcitrant in the future, we can treat them earlier and more aggressively to block the progression of the disease.
MATERIALS AND METHODS
Subjects and material collection
The study subjects consisted of 281 AD patients who had visited the outpatient clinic of the Department of Dermatology, Wonju Severance Christian Hospital from December 2008 to September 2017. All of the AD patients were diagnosed by dermatologists. Basic information such as age, sex, onset age or duration of AD, and family and personal history of AD, allergic rhinitis, and asthma were collected along with initial eczema area severity index (EASI) score. Serum total IgE level (ADVIA Centaur Total IgE; Siemens, New York, NY, USA) and multiple allergen simultaneous test (MAST) (AdvanSure AlloScreen; LG Chemistry, Seoul, Korea) were performed and serum IgE level for Dermatophagoides farinae and Dermatophagoides pteronyssinus was recorded. Intrinsic AD was defined as the subjects in which total serum IgE was less than 200 IU/mL and negative in MAST test. Whereas, extrinsic AD was defined as patients in which the total IgE is equal or more than 200 IU/mL, or MAST test was positive for at least one allergen. This study was approved by the Yonsei University Wonju Campus Institutional Review Board and was performed in accordance with their guidelines.
AD treatment, evaluation of treatment response, and subject groups
All AD patients were instructed to continue using proper moisturizer and avoid individual aggravating and/or trigger factors. They were treated with one or more of the followings for the indicated: topical corticosteroids, topical calcineurin inhibitors, systemic antihistamines, gamma linoleic acid, and topical or systemic antibiotics. Treatment response was assessed by patient follow up to evaluate factors including pruritus score by visual analog scale and EASI score. If the treatment response was not considered satisfactory and there were no inhibitory factors such as underlying disease or abnormal laboratory test results, systemic immunomodulators such as cyclosporine, methotrexate, or azathioprine were used. If the EASI score was not decreased by 50% after 4 weeks of one or more of systemic immunomodulator use, the patient was classified as a ‘recalcitrant’ AD patient. If a patient did not use systemic immunomodulators or showed a satisfactory treatment response within 4 weeks of systemic immunomodulator treatment, the patient was designated as a ‘treatable’ AD patient.
Candidate gene variations selection, genomic DNA extraction, multiplex polymerase chain reaction (PCR) amplification, and sequence analysis
By reviewing previous studies on the relationships between AD and SNPs, candidate genes that are highly related to AD among East Asian people including Koreans were identified. Based on the results of previous genetic studies in Koreans and our previous large-scaled study,13 AD-associated candidate genes such as 6 barrier function-related gene variations (FLG 3321delA, FLG K4022X, KLK7, SPINK 1156, SPINK 1188, and SPINK 2475) and 6 immune response-related gene variations (DEFB1 [rs5743399], KDR [rs2305948], IL-5RA [rs334809], IL-9 [rs31563], and IL-12RB1 [rs393548 and rs436857]) were selected. Afterward, the presence and frequency of these gene variations were compared between the treatable and recalcitrant AD groups.
Specific primer sequences for KLK7, SPINK5, FLG, DEFB1, KDR, IL-5RA, IL-9, and IL-12RB1 were obtained using data from the GenBank database of the National Center for Biotechnology Information (http://www.ncbi.nlm.nih.gov). Using these primers, PCR was performed on genomic DNA extracted from blood samples of the subjects, utilizing the QIAamp DNA Mini Kit (Qiagen GmbH, Hilden, Germany). The amplification mixture for the KLK7, SPINK5, FLG, DEFB1, KDR, IL-5RA, IL-9, and IL-12RB1 genes contained 1X primer-mix, 2X PCR premix (Genetbio, Daejeon, Korea), 2 mM of MgCl2, 250 µm dNTP, and 10 ng of genomic DNA in a final volume of 50 µL. Multiplex PCR was then performed using this amplification mixture, followed by direct sequencing of both strands of PCR product using the ABI 3100 Genetic Analyzer (Applied Biosystems, Foster City, CA, USA) at COSMO Genetech Co. LTD. (Seoul, Korea). Subsequent sequence alignment was performed using multiple sequence alignment programs (http://bioinfo.genopole-toulouse.prd.fr).
Reverse blot hybridization assay (REBA)
The genus-specific oligonucleotide probes of KLK7, SPINK5, FLG, DEFB1, KDR, IL-5RA, IL-9, and IL-12RB1 were designed using sequence data from the NCBI database (Bethesda, MD, USA), followed by a BLAST search (National Center for Biotechnology Information of the National Library of Medicine website) to confirm the sequence homology for each probe. Thereafter, a REBA membrane that can detect a total of 26 probes including 13 wild-type (WT) and 13 mutant-type (MT) probes was designed to detect KLK7, SPINK5, FLG, DEFB1, KDR, IL-5RA, IL-9, and IL-12RB1 and one control probe using colorimetric signal detection. The REBA was performed as follows: First, 20 µL of each PCR product was mixed with an equal volume of denaturation solution (0.2 N NaOH, 0.2 mM ethylenediaminetetraacetic acid) and incubated at room temperature (RT) for 5 minutes. Next, the denatured PCR products were diluted with 960 µL of 2X SSPE/0.1% SDS. The REBA membrane strips were then placed on a Minitray (Bio-rad, Hercules, CA, USA) and incubated with 2X SSPE/0.1% SDS for 5 minutes. After residual fluid in the slot was removed, the slots were filled with denatured single-stranded PCR products. The PCR products were incubated at 55℃ for 30 minutes, washed twice with 2X SSPE/0.5% SDS at 62℃ for 10 minutes, and incubated in 1:2,000 diluted streptavidin-conjugated alkaline phosphatase (Roche Diagnostics, Mannheim, Germany) with 2X SSPE/0.5% SDS for 30 minutes at RT. The hybridized amplicons were detected colorimetrically by incubating the strip in 1:50 diluted nitro blue tetrazolium chloride/5-bromo-4-chloro-3-indolyl phosphate, toluidine salt (NBT/BCIP, Roche Diagnostics) in 67% dimethyl sulfoxide (v/v) with tris-buffered saline (pH 9.5) for 5–10 minutes. The presence of the MT and WT of each probe was confirmed by analyzing the band pattern. All experimental procedures were performed in duplicate.
Statistical analysis
The two-sample t test, χ2 test or Fisher's exact test, and multivariable logistic regression were performed as appropriate. Differences in the genetic variation of KLK7, SPINK5, FLG, DEFB1, KDR, and IL-5RA, IL-9, and IL-12RB1 were identified through the χ2 test or Fisher's exact test. In calculating adjusted odds ratio (OR), multivariate logistic regression was performed. The multiple logistic regression model included demographic data, the frequency of each SNPs in each AD group, and the subjects' age as covariates.
All statistical analyses were conducted using SAS version 9.4 (SAS Inc., Cary, NC, USA), and a P value <0.05 was considered statistically significant.
RESULTS
Subject characteristics
In total, 249 treatable AD patients (88.6%) and 32 recalcitrant AD patients (11.4%) were recruited. The mean age of recalcitrant AD patients was higher than that of treatable AD patients (31.3 vs 15.3 years, respectively). The subjects with self-reported history of AD had higher odds of recalcitrant AD compared to the subjects (age-adjusted OR, 12.37; 95% confidence interval [CI], 3.17–62.34). Initial EASI score was higher in recalcitrant AD group (8.6 vs 19.5, respectively) and had positive association with recalcitrant AD (age-adjusted OR, 1.13 per 1 EASI; 95% CI, 1.09–1.19 per 1 EASI). Clinically, nummular eczema was more frequently found in recalcitrant AD group (11.2% vs 34.4%, respectively), and had higher odds of recalcitrant AD (aged-adjusted OR, 2.76; 95% CI, 1.08–6.65). In comparing serologic markers, total IgE levels were higher in recalcitrant AD than in treatable AD patients (573.5 vs 1,384.3 IU/mL, respectively). The subjects with total IgE ≥1,000 IU/mL had higher odds of recalcitrant AD (age-adjusted OR, 5.55; 95% CI, 1.58–25.99). House dust mite (HDM) specific IgE ≥1,000 IU/mL was strongly associated with recalcitrant AD (D. farinae [age-adjusted OR, 11.16; 95% CI, 3.94–37.68] and D. pteronyssinus [age-adjusted OR, 10.82; 95% CI, 4.04–33.71], respectively). Detailed characteristics of the subjects are summarized in Table 1.
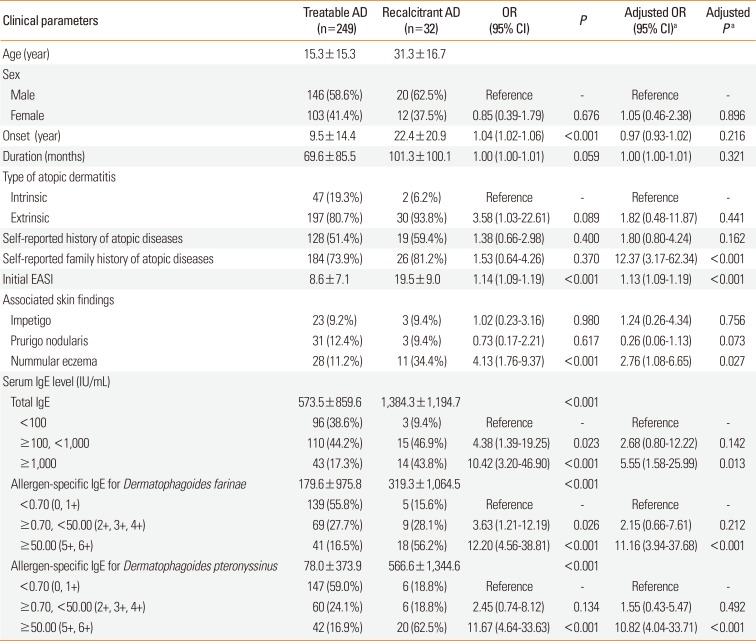
Table 1
Comparisons of clinical characteristics between “Treatable AD” and “Recalcitrant AD”
![]()
Differences in barrier function-related gene variation frequencies between treatable and recalcitrant AD patients
The frequency of Hetero, homozygous mutation (Homo), and WT in each barrier function-related genes was compared between the treatable and the recalcitrant AD group. Hetero in KLK7 were more frequent in the recalcitrant AD group than in treatable AD group but without statistical significance (68.8% vs 50.6%, respectively; age-adjusted OR of recalcitrant AD, 2.38; 95% CI, 0.97–6.51). Similarly, Homo in SPINK C1188T was more frequent in the recalcitrant AD group than in treatable AD group but without statistical significance (31.2% vs 20.9%, respectively; age-adjusted OR of recalcitrant AD, 2.53; 95% CI, 0.84–8.18). In our study, no Homo of FLG gene was observed, and Hetero of FLG 3321delA and FLG K4022X accounted for ≤10% both in treatable and recalcitrant AD group. No evident differences between recalcitrant AD and treatable AD group was observed in the frequency of Hetero of FLG (Table 2).
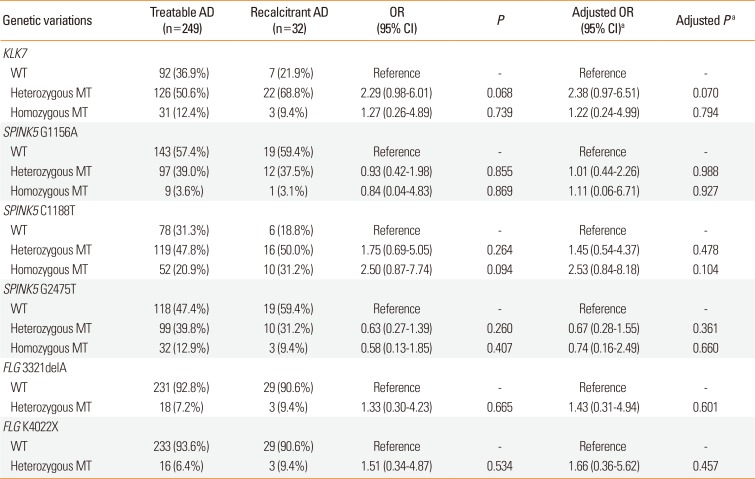
Table 2
Comparisons of barrier-related genetic variations between “treatable AD” and “recalcitrant AD”
![]()
Differences in immune reaction-related gene variation frequencies between treatable and recalcitrant AD patients
A slightly different tendency was observed in immune reaction- related gene variations. First, Homo in DEFB1 C2266T was more frequent in recalcitrant AD group than in treatable AD group (25.0% vs 17.3%, respectively). Although tendency to recalcitrant AD group was observed, there was no statistical significance (age-adjusted OR, 0.92; 95% CI, 0.31–2.65). Instead, Hetero of DEFB1 C2266T was less frequent in recalcitrant AD than in treatable AD (40.6% vs 61.4%, respectively; age-adjusted OR of recalcitrant AD, 0.42; 95% CI, 0.13–0.89). Likewise, Hetero of KDR was less frequent in recalcitrant AD than in treatable AD but without statistical significance (18.8% vs 22.1%, respectively; age-adjusted OR of statistical significance, 0.86; 95% CI, 0.30–2.19).
Hetero of IL-5RA and Homo of IL-12RB1b was more frequent in recalcitrant AD. In contrary, Hetero of IL-I12RB1a was less frequent in recalcitrant AD than in treatable AD. Hetero of IL-9 showed no difference between treatable and recalcitrant AD group. And all these differences had no statistical significance (Table 3).
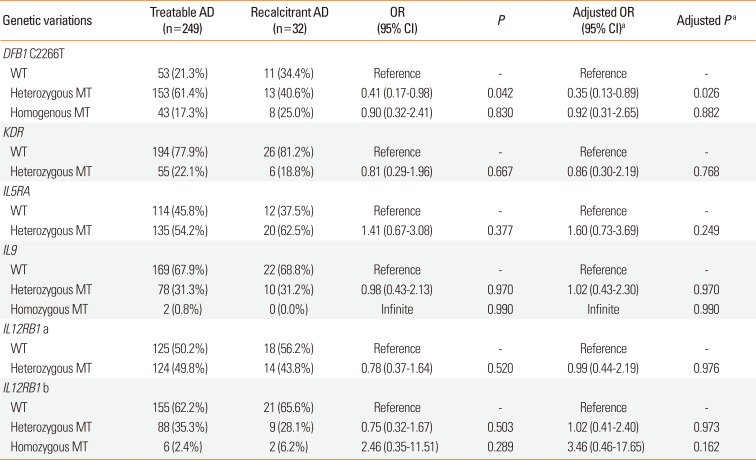
Table 3
Comparisons of immune-response related genetic variations between “treatable AD” and “recalcitrant AD”
![]()
DISCUSSION
The development of AD is influenced by variations in not only immune response-related genes such as DEFB, TNFA, KDR, FCER1A and IL-4, 5, 9, 10, 12, 13, and 18, but also in barrier function- related genes such as FLG, SPINK5, and KLK7.46151618192122242829 Previous studies have demonstrated a link between AD and these diverse genetic variations, so we aimed to further investigate whether this genetic diversity affects the treatment response of AD.
Filaggrin is incorporated into the lipid envelope of the stratum corneum and plays an important role in water retention of the skin through its degradation into free amino acids.30 The frequency of FLG mutations varies according to race. While up to 50% of Caucasian patients with severe eczema have at least one FLG mutation,31
FLG mutations were observed in less than 10% of Koreans with AD, among which pK4022X (5%) and c3321delA (1.8%) were the most common.32 In addition, according to our previous report, FLG 3321delA SNP was found to be significantly more frequent in patients with moderate to severe AD than in normal controls.13 In this study, Hetero in FLG 3321delA and FLG K4022X was slightly more frequent in recalcitrant AD patients than in treatable AD patients. However, these differences did not show statistical significance, and also the fraction of Hetero of FLG was less than 10% in both the recalcitrant AD and treatable AD groups, as indicated in previous studies. Considering the relatively small subject size of our study, it is possible that the FLG mutation in Koreans might re-emerge in future large-scale studies. Nevertheless, given that previous studies on the correlation between FLG mutations and AD in Koreans and the results of our study, the effect of FLG mutations is likely to be smaller in Koreans than in Northwestern Europeans, both in the occurrence and the treatment response of AD.
KLK7 encodes the stratum corneum chymotryptic enzyme (SCCE) and is involved in desquamation and maintenance of epidermal homeostasis.33 Small mutations such as an AACC insertion in the KLK7 gene region can significantly increase the incidence of AD.634
SPINK5 encodes a lympho-epithelial Kazal-type-related inhibitor (LETKI), which affects the action of SCCE. A loss of function mutation in SPINK5 affects keratinocyte cornification and is significantly correlated with atopic diseases such as AD, allergic rhinitis, and asthma and elevation in serum IgE and eosinophils.35 In this study, there was no statistically significant difference in the frequency of Homo in KLK7, SPINK 1188, and SPINK 2475 between the treatable and recalcitrant AD groups. However, according to our previous study in Korean AD patients, Hetero in SPINK5 1156 and Homo in SPINK5 2475 were more frequent in the AD group than in the control group.13 Also, it has been reported that KLK7 dysfunctions and SPINK5 variations are associated with AD in various prior studies predominantly examining Caucasians. When interpreting our findings based on these prior studies, it is possible that Homo of KLK7 and SPINK5 clearly affects onset of AD, but its association with AD treatment response is relatively small or insignificant in East Asians, especially Koreans.
Defensin is an AMP that plays a role in innate immunity. The lack of β-defensin has been found to trigger the colonization of bacteria or candida species in patients with AD.1635 Vascular endothelial growth factor (VEGF) is involved in epithelial cell proliferation and blood vessel formation, which is associated with its major receptor, VEGF receptor 2. KDR plays an important role in maintaining the high association rate between VEGF and VEGF receptors and is closely related to antigen sensitization and the manifestations of atopic disease.14 It is also known that the V297I mutation in KDR is closely related to atopic disease.36 Contrary results have been reported for the role of DEFB1 in AD. In 2009, Kim et al.15 reported a significant correlation between Korean AD and the haplotype DEFB1. However, in a study in northeast Brazilians in 2010, Segat et al.37 reported that there is a high likelihood that there is no significant role of DEFB1 in AD. Meanwhile, Prado Montes de Oca and Li38 reported that variations in DEFB1 might be related to the ethnic susceptibility of AD. In addition, according to our previous report from Korean AD patients, Hetero in DEFB1 (rs5743399) and KDR were found to be more common in patients with AD than in the normal control group.13 According to the result of our study, Hetero of DEFB1 is less frequent in recalcitrant AD than in treatable AD. These results imply that the Hetero of DEFB1 is a factor affecting the onset of AD, but paradoxically, the presence of the Hetero of DEFB1 might be a good prognostic factor in terms of AD treatment. Given the relatively small size of the recalcitrant AD group in this study and the absence of specific differences in the KDR genes, further large-scaled studies of this area are needed to demonstrate a significant correlation between AD and DEFB1/KDR variations in East Asian populations, especially in Koreans.
According to our previous report (Supplementary Table),13 Hetero in IL-5RA (rs334809), Hetero in IL-9 (rs31563), and 2 Hetero in IL-12RB1 (rs393548, rs436857) were more frequent in the AD group than in the control group. These results are consistent with prior studies on the relevance between ILs and AD.21 However, IL-5RA, IL-9, IL-12RB1 showed no significant difference between recalcitrant AD and treatable AD. This result implies that Hetero in IL-5RA, IL-9, IL-12RB1 is closely related to the occurrence of AD, but suggests that the treatment response of AD is unlikely to be deeply involved with these SNPs.
Our results indicate that recalcitrant AD patients are older than treatable AD, and especially the onset age is higher in recalcitrant AD. However, this difference went statistically insignificant when age was adjusted. Other than age and onset age, clinical markers such as family history of atopic diseases, Initial EASI score, associated nummular eczema, total IgE ≥1,000, and allergen-specific IgE for HDMs are significantly associated to the refractory treatment response, even after age-adjustment. Especially, OR of recalcitrant AD was 5.55 when total serum IgE ≥1,000, and which was greater than 10 when allergen-specific IgE for HDM was ≥1,000. When we associate these results with previous studies and our results of genotype comparisons, although it is evident that variations in barrier-associated and immune-related genes have a clear impact on the occurrence of AD, clinical finding is more likely to be a more important prognostic factor in predicting the response of AD, to these genetic variants.
Another interesting finding is that the OR of recalcitrant AD was significantly increased by 12 or more when there was a self-reported family history of atopic diseases. We collected history of atopic diseases such as allergic rhinitis, asthma, and AD during the collecting of research data. The association of AD with these diseases is well known. Although our treatable AD group and recalcitrant AD group are not affluent in numbers, and there is a limitation in that there was a discrepancy in the number of treatable and recalcitrant AD group, it could be concluded that AD will show a poor response to treatment when there is a history of such atopic diseases. Surely, in comparing the genotypes between the recalcitrant and the treatable AD, the possibility of influence of these phenotypic differences could not be completely ruled out. However, as treatable AD was defined as patients who showed clinical improvement in the use of cyclosporin, methotrexate (MTX), or azathioprine for ≤4 weeks of treatment, recalcitrant AD was defined as patients who did not show ≥50% clinical improvement after treatment with cyclosporin, MTX, and azathioprine for ≥4 weeks. Thus, the difference in these phenotypes cannot be a confounding variable, and as genetic variation is not a factor that depends on age or disease duration, it is less likely that it will impair statistical significance of differences in genotype of treatable and recalcitrant AD.
One of the major limitations of this study is that the number of recalcitrant AD group is limited to completely exclude out the genetic influence on recalcitrant disease. However, AD subjects with treatment-resistant and refractory conditions were relatively limited, compared to other eczematous dermatoses. Thus, a definitive conclusion for genetic influence on recalcitrant disease would require a larger and further studies with long-term cumulative data.
We found that variations in representative barrier-associated and immune response-related genes were not likely to significantly affect the AD treatment response. Although previous studies have shown that the effect of these variation on AD occurrence is clear, the likelihood that this effect will affect the AD treatment response is not high at this time. Rather, clinical findings such as initial EASI score, serum IgE level, and family history of atopic diseases were found to have a more significant impact on adverse treatment response. In a variety of preceding studies, variations in immune dysregulation-related genes not included in our study are also found. However, this study is distinguished in that it focused on the association between long-term treatment response and gene variation rather than AD severity at the time of diagnosis. If we can predict whether AD patients will become recalcitrant in the future, we will be able to treat them earlier and more aggressively to block the progression of the disease.
 XML Download
XML Download