

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Asthma is a chronic inflammatory airway disease characterized by airway hyperresponsiveness (AHR), inflammation, and remodeling.1 The prevalence of asthma has been increasing over the past few decades, and its economic burden is substantial, especially in uncontrolled asthma.2345 It has become a major public health problem. Understanding the precise pathophysiology of asthma is important to achieve optimal management.
The transient receptor potential vanilloid 1 (TRPV1) channel is a non-selective calcium ion (Ca2+) channel that is expressed in various cell types, including sensory neurons, epithelial cells, and smooth muscle cells.67 It is stimulated by various stimuli, including noxious chemicals, low pH, hot temperature, and endogenous mediators.89 Many studies have sought to reveal the role of the TRPV1 channel in airway diseases, with an emphasis on chronic cough. TRPV1 expression in the airway nerves is increased in chronic cough patients, and TRPV1 antagonists may have potential value as antitussive drugs.101112 Mucus hypersecretion and airway inflammation may also be associated with TRPV1 sensitization.13
The role of TRPV1 in the pathophysiology of asthma has attracted attention. TRPV1 expression is increased in the airway epithelium of asthmatic patients and is more prominent in severe, uncontrolled disease.14 However, the roles of TRPV1 and the effects of TRPV1 antagonism on airway inflammation in animal models are debatable. Rehman et al.15 reported that inhibition of TRPV1 reduced AHR and airway remodeling in interleukin (IL)-13-induced asthma model in BALB/c mice. However, Caceres et al.16 induced an acute asthma murine model in genetically silenced C57BL/6 mice using ovalbumin (OVA) and obtained negative results. The results of TRPV1 blocking are diverse in different experimental materials and study settings.
In addition to the contrary results of TRPV1 antagonism in diverse murine asthma models, there was also lack of studies performed in a chronic asthma model compared to acute model. Chronic asthma model may demonstrate better asthma features, especially airway remodeling, which is important in human asthma pathophysiology.
In this study, we investigated the role of TRPV1 in the airway of a murine model of chronic asthma. We also examined whether TRPV1 suppression by inhalation of antagomir, a small interfering RNA (siRNA) directed toward TPRV1, may alleviate pathologic manifestation of chronic asthma compared to the well-known TRPV1 inhibitor capsazepine.
MATERIALS AND METHODS
Animals and experimental design
Six-week-old female BALB/c mice (Orient Bio Inc., Seongnam, Korea) were randomly allocated to the following groups: 1) control, 2) OVA challenge, 3) OVA challenge plus capsazepine, or 4) OVA plus TRPV1 siRNA.
Sensitization and antigen challenge protocol
Sensitization and antigen challenge with OVA were performed as previously described.1718 Mice were immunized by subcutaneously injecting 25 µg of OVA (Grade V; Sigma-Aldrich, St. Louis, MO, USA) absorbed to 1 mg of aluminum hydroxide (Aldrich, Milwaukee, WI, USA) in 200 µL of phosphate-buffered saline (PBS). Subcutaneous injections were administered on days 0, 7, 14, and 21, followed by intranasal OVA challenge (20/50 µL in PBS) performed on days 33 and 35. Subsequently, intranasal OVA challenges were repeated twice per week for 3 months. Age- and gender-matched control mice were treated equally with PBS. All procedures were performed while mice were anesthetized using isoflurane (Vedco, St. Joseph, MO, USA). Mice were sacrificed 24 hours after the final intranasal OVA challenge, and bronchoalveolar lavage (BAL) fluid and lung tissues were obtained for analysis.
All animal procedures were performed in accordance with Laboratory Animal Welfare Act, the Guide for the Care and Use of Laboratory Animals, and the Guidelines and Policies for Rodent Experiments provided by Institutional Animal Care and Use Committee at the School of Medicine, The Catholic University of Korea (approval number: CUMC-2015-0194-04).
Administration of capsazepine and TRPV1 siRNA
Capsazepine (Cayman, Ann Arbor, MI, USA) was given 50 µg once daily for 3 months by intraperitoneal injection starting on day 38. TRPV1 siRNA (Bioneer, Daejeon, Korea) was also administrated intranasally 50 µg 2 times per week once per day beginning on day 38 for 3 months, during OVA challenge. The control mice were treated identically with normal saline.
Measurement of AHR
AHR to methacholine (Mch)(Sigma-Aldrich) was assessed 24 hours after the final OVA challenge with the flexiVent system (SCIREQ, Montreal, Canada) as previously described.19 Briefly, mice were anesthetized with an intraperitoneal administration of a 1:4 mixture of rompun and zoletil. The trachea was exposed and cannulated to connect it with a computer-controlled small-animal ventilator. Ventilation was applied with a tidal volume of 10 mL/kg at a frequency of 150 breaths/min and a positive end-expiratory pressure of 2 cm H2O, which was close to the mean lung volume of mouse spontaneous breathing. Each mouse was challenged with PBS control, followed by Mch aerosol with increasing concentrations (6.25, 12.5, 25, and 50 mg/mL). Changes in airway resistance with increasing concentrations of inhaled Mch were measured.
BAL
Mice were sacrificed by CO2 asphyxiation after measurement of AHR. The trachea was exposed and cannulated with a silicone tube attached to a 23-gauge needle on a 1-mL tuberculin syringe. BAL was performed by instillation of 0.8 mL of sterile PBS through the trachea into the lung. The total cell counts in BAL fluid were analyzed using a LUNA™ Automated Cell Counter (Logos Biosystems, Inc., Annandale, VA, USA).
The BAL fluid was cytospun at 2,000 rpm for 7 minutes, placed on microscope slides, and stained with Diff-Quick (Sysmax, Kobe, Japan). The percentages of macrophages, eosinophils, lymphocytes, and neutrophils in the BAL fluid were calculated by counting 500 leukocytes on randomly selected areas of the slide using light microscopy. Supernatants were stored at −70℃.
Lung tissue histopathology
After BAL was performed, the mouse lungs were inflated, fixed in 4% paraformaldehyde for 24 hours, and embedded in paraffin using a standard protocol. Sections were cut 4-µm thick using a microtome and stained with hematoxylin and eosin (H&E). Paraffin-embedded tissues were also sectioned and the 5- to 6-µm thick sections were stained with periodic acid-Schiff (PAS) to distinguish goblet cells in the epithelium. Goblet cell hyperplasia was quantified as previously described.20 The pathological changes were evaluated according to a modified 5-point scoring system (grades 0–4) based on the percentage of goblet cells in the epithelium: grade 0 (no goblet cells), grade 1 (<25%), grade 2 (25%–50%), grade 3 (51%–75%), and grade 4 (>75%). The mean goblet cell hyperplasia score was then calculated for each mouse.
Enzyme-linked immunosorbent assay (ELISA)
The concentrations of IL-4, IL-5, and IL-13 were measured in BAL fluid. Concentrations of IL-17E (IL-25), IL-33, and thymic stromal lymphopoietin (TSLP) were measured with lung homogenate with an ELISA kit (R&D Systems, Minneapolis, MN, USA). The assay was performed according to the manufacturer's protocol.
Immunohistochemistry
Six-µm-thick lung sections from each paraffin block were deparaffinized with xylene and rehydrated in ethanol. For immunohistochemical detection of α-smooth muscle actin (α-SMA) and TRPV1, the lung sections were incubated overnight at 4℃ with a primary monoclonal antibody against α-SMA (titer 1:25; Abcam, Cambridge, MA, USA) and TRPV1 (titer 1:100; Abcam), or mouse serum as a negative control instead of the primary antibody. Immunoreactivity was detected by sequential incubations of lung sections with a biotinylated secondary antibody, followed by peroxidase reagent and the 3,3′-diaminobenzidine (DAB) chromogen (Invitrogen, Carlsbad, CA, USA). The area in each paraffin-embedded lung immunostained by α-SMA was outlined and quantified using a light microscope attached to an image analysis system (BX50; Olympus, Tokyo, Japan). The results were expressed as the immunostained area of the bronchiolar basement membrane (internal diameter 150–200 µm). At least 10 bronchioles were counted in each slide.
Hydroxyproline analysis
Lung tissue (60 mg) from each mouse was used for the hydroxyproline assay with hydroxyproline colorimetric assay kit (BioVision, Milpitas, CA, USA) according to the manufacturer's instructions. Hydroxyproline concentrations were calculated from a standard curve of hydroxyproline.
Data analysis
The results from each group were analyzed by analysis of variance (ANOVA) and the nonparametric Kruskal-Wallis test. All statistical analyses were performed using Graph-Pad Prism for Windows software (ver. 5.00; GraphPad Software, San Diego, CA, USA). A P value of <0.05 was considered statistically significant. All results are expressed as mean±standard error of the mean (SEM).
RESULTS
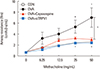
Inhibitory effects of TRPV1 antagonists on AHR
The OVA challenge group showed increased AHR compared to the control group at Mch doses of 25 and 50 mg/dL. The OVA plus capsazepine group displayed a remarkable decrease in airway resistance at 50 mg/mL Mch. Airway resistance was significantly decreased in mice treated with TRPV1 siRNA at Mch doses of 25 and 50 mg/dL (Fig. 1).
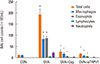
Effects of TRPV1 inhibition on eosinophilic airway inflammation and the level of cytokines
Three months of OVA challenge induced a significant increase in number of total cells, macrophages, and eosinophils in the BAL fluid. Treatment with either capsazepine or TRPV1 siRNA significantly reduced the number of total cells, macrophages, and eosinophils in BAL fluid (Fig. 2). Three months of OVA challenge significantly increased the levels of type 2 T helper (Th2)-associated cytokines (IL-3, IL-5, and IL-13) in BAL fluid (Fig. 3A). Mice treated with capsazepine displayed significant decreases in IL-4, IL-5, and IL-13. Treatment with TRPV1 siRNA significantly decreased IL-4 and IL-5. We evaluated the epithelial cell-derived cytokines IL-17E (IL-25), IL-33, and TSLP in the lung homogenate. The OVA challenge group showed increases in all the 3 cytokines compared to the control group (Fig. 3B). Mice treated with either capsazepine or TRPV1 siRNA showed significantly decreased levels of IL-33 and TSLP. However, IL-17E level was decreased in both group, without statistical significance.
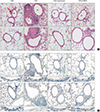
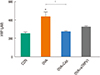
Effects of TRPV1 inhibitors on expression of TRPV1 receptors and airway remodeling
Histopathology of H&E-stained sections revealed increased infiltration of subepithelial, peribronchial, and perivascular inflammatory cells, including eosinophils, in the OVA challenge group compared to the control group. In contrast, treatment with either capsazepine or TRPV1 siRNA reduced the inflammatory cell recruitment (Fig. 4A). Immunostaining of TRPV1 showed that chronic OVA challenge increased the area of TRPV1 immunostaining compared to the control group. This effect was alleviated by capsazepine or TRPV1 (Fig. 4B). PAS-stained sections revealed prominent goblet cell hyperplasia in the OVA challenge group. The hyperplasia was attenuated by both treatments (Fig. 5A). The pathologic score was significantly higher in the OVA group compared to both treatment groups (Fig. 5B). Quantification of peribronchial α-SMA immunostaining was performed by imaging (Fig. 6). Chronic OVA challenge induced an increase in the area of peribronchial α-SMA compared to the control group. Treatment with capsazepine or TRPV1 siRNA reduced the area of peribronchial α-SMA. Pulmonary fibrosis was evaluated by measuring the total lung collagen level by using hydroxyproline analysis. Chronic OVA challenge for 3 months resulted in a significant increase in hydroxyproline content (Fig. 7). Capsazepine treatment significantly decreased the level of hydroxyproline compared to the OVA challenge group. TRPV1 siRNA treatment decreased the hydroxyproline level, without statistical significance.
DISCUSSION
This study revealed that expression of the TRPV1 receptor was significantly elevated in OVA challenged asthmatic lungs and that inhalation of TRPV1 antagomir effectively suppressed eosinophilic airway inflammation and remodeling. The underlying mechanisms of these effects may be associated with suppression of epithelial-derived cytokines, including TSLP, IL-33, and IL-25. This is the first report that the inhibition of the TRPV1 receptor by TRPV1 antagomir inhalation has significant anti-asthmatic effects that are comparable to the established TRPV1 antagonist, capsazepine.
The 3-month chronic OVA-challenged model is unique in that it results in features that are more similar to human asthma than the acute model, which features airway remodeling, including airway smooth muscle cell (ASMC) proliferation, goblet cell hyperplasia, and collagen deposition. The present results are important since, to our knowledge, no prior study has explored the role of TRPV1 in a chronic allergic asthma animal model.
Asthma is an allergic inflammatory airway disease in which pathogenesis involves various inflammatory cells and mediators. The most important cellular pathway of asthma starts with recognition of allergen by dendritic cells in airway epithelium and submucosa.21 As dendritic cells function as antigen-presenting cells to Th2 lymphocytes, Th2 cytokines are released. This release induces eosinophils and mast cells to initiate allergic inflammation.22 In this process, bronchial epithelial cells play an important role in asthma pathophysiology. They function as a mechanical barrier from allergens and other toxic materials through tight junctions, and also release cytokines—including IL-25, IL-33, TSLP, and granulocyte macrophage-colony stimulating factor—or endogenous danger signals in response to various allergens to modulate allergic inflammation.23
The TRPV1 channel is expressed in both sensory neurons and non-neuronal cells, including bronchial epithelium, smooth muscle cells, mast cells, and dendritic cells.811142425 Consistent with a previous study, we identified increased TRPV1 expression of epithelial cells by immunochemical staining in the murine asthma model. Activation of TRPV1 in bronchial epithelial cells stimulates the release of proinflammatory mediators, including ILs, prostaglandin E2, nerve growth factor, and tumor necrosis factor-α, to promote airway inflammation and airway hypersensitivity.62627
Recent studies have demonstrated that TRPV1 is important in the pathway of airway inflammation and remodeling, which is a key pathologic feature of asthma.28293031 Earlier studies reported that inhibition of TRPV1 reduced airway constriction using capsazepine and in a knockout animal model. Ellis and Undem32 revealed that capsazepine inhibits capsaicin-sensitive receptors that attenuate airway constriction. Furthermore, Watanabe et al.33 demonstrated that loss of TRPV1-positive axon results in alleviation of bronchoconstriction. Also, there are some studies that explored the role of TRPV1 receptor in asthma animal models. The findings have been inconsistent. Rehman et al.15 observed that TRPV1 siRNA treatment in acute asthma murine model significantly alleviates asthma features. Also, Baker et al.25 reveal attenuation of asthma features in genetically silenced TRPV1, although the results did not reach statistical significance. Caceres et al.16 reported that TRPV1 knockout mice showed no difference, compared to the wild type, in AHR or airway inflammation in an OVA-challenged asthma model. Raemdonck et al.34 demonstrated a similar result that asthma was not attenuated by the TRPV1 blockers JNJ-17203212 and capsazepine in an OVA-induced murine asthma model. Moreover, Mori et al.35 suggested that TRPV1 gene knockout mice intensified AHR and airway inflammation compared to the wild type in an asthma model, which is the exactly the opposite result of our study. These conflicting results may reflect the mouse strains used; the prior study involved C57BL/6 mice. Distribution of capsaicin receptors varies with strain, and there may be less TRPV1 expression in C57BL/6 mice than in BALB/c mice.1536
Currently, TRPV1 expression was significantly elevated in lung tissues of the OVA-induced asthma model. In addition, we confirmed that capsazepine (TRPV1 antagonist) and TRPV1 siRNA (antagomir) inhalation resulted in the attenuation of AHR and inflammation. Antagomir inhalation for 3 months showed noninferior effects compared to capsazepine.
In this study, we found that anti-inflammatory effects on the asthmatic airway may be associated with decreased expression of epithelial-derived cytokines and Th2 cytokines by TRPV1 blocking. Epithelial cell-derived cytokines, such as TSLP, IL-33, and IL-25, are important regulators of Th2 cytokine associated inflammation.3738 IL-33 and IL-25 activate Th2 lymphocytes and other cells, which mediate allergic reactions and activate TSLP production. TSLP is important in the activation of dendritic cells and naïve T cells. In our study, the levels of IL-33 and TSLP were significantly decreased after TRPV1 blocking. Also, IL-25 displayed a decreasing trend, which did not reach statistical significance. These results imply that TRPV1 channel may play a crucial role in bronchial epithelial cells after chronic OVA challenge and affect Th2-mediated inflammation by regulating epithelial cell-derived cytokine expression. However, the limitation of our study is that exact mechanism underlying down-regulation of IL-33 and IL-25 by TRPV1 blocking have not been revealed. Further studies are needed to reveal the role of TRPV1 on airway epithelial cells and other inflammatory cells, including in vitro experiment using the epithelial cell line.
Airway remodeling is an essential pathophysiologic feature of chronic bronchial asthma.39 Currently, TRPV1 antagonism alleviated airway remodeling by reducing airway smooth muscle thickening and collagen deposition. Until now, the effect of TRPV1 inhibition on airway remodeling has not been fully understood. One of the most important features of airway remodeling is ASMC hypertrophy and hyperplasia.22 Zhao et al.28 revealed that the TRPV1 channel is overexpressed and activated in ASMCs of asthmatic rats. Treatment with capsaicin has been reported to enhance ASMC proliferation and decrease apoptosis, whereas capsazepine did in an opposite manner. ASMC hypertrophy and hyperplasia are important features of airway remodeling. Thus, the TRPV1 channel in ASMCs may play a crucial role in airway remodeling in asthma. Furthermore, the accumulation of matrix proteins, such as collagen fibers, is also responsible for airway thickening in chronic asthma.22 A previous study revealed that, although TRPV1 channel is not generally expressed in airway fibroblasts, it is expressed significantly in inflammatory conditions induced by tumor necrosis factor-α, lipopolysaccharide, and IL-1α.26 This result suggests that bronchial fibroblasts may be activated to synthesize collagen fibers in inflammatory conditions, such as bronchial asthma, via the TRPV1 channel. Further studies are needed to understand the exact mechanism of TRPV1 in airway remodeling.
In conclusion, blocking the TRPV1 pathway by capsazepine or TRPV1 siRNA inhalation attenuates OVA-mediated asthma features, including allergic inflammation, AHR, and airway remodeling. The TRPV1 antagonist delivered via nasal inhalation might have therapeutic potential in the treatment of bronchial asthma.



 XML Download
XML Download