

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
The cerebellum, located in the posterior cranial fossa, is a major motor structure of the brain. It controls motor-related functions, such as maintaining balance and posture, and motor learning including coordination of movements through complex regulatory and feedback mechanisms. Cerebellar lesions cause dysmetria, ataxia, and intention tremors.123 The cerebellum is also associated with the control of cognition and emotion, and patients with cerebellar dysfunction experience cognitive and affective symptoms.456
Mediolaterally, the cerebellum consists of the midline vermis and two cerebellar hemispheres. 7 In the anterior-posterior division, it can be divided into anterior, posterior, and flocculonodular lobes, which are further separated into ten lobules that are designated as I–X.8 The anterior lobe (comprising lobules I–V) is divided from the posterior lobe by the primary fissure, and the posterior lobe (comprising lobules VI–IX) is divided from the flocculonodular lobe (lobule X) by the posterolateral fissure.9
The medial zone consists of the vermis and nuclei fastigii (NF),7 which is primarily associated with the control of posture and locomotion. Lesions in the vermis in humans and inactivation of the NF in monkeys induce deficits in sitting, standing, and walking.101112 The nucleus interpositus (NI) and those portions of the paravermal cortex that project to these nuclei form the intermediate zone.7 Transitory inactivation of the NI and the adjacent portion of the dentate nucleus (DN) in monkeys results in tremor11 and deficits in reaching and grasping motions.131415 The lateral cerebellar zone consists of the cerebellar hemispheres and most of the DN.7 Its anterior portion participates in motor control, while the posterolateral portion is involved in motor planning, language production, and cognitive processes such as memory function.161718
Different regions of the cerebellar cortex form microcomplexes that process information delivered by the inputs of each microcomplex' equally.319 This means that different regions of the cerebellum are involved depending on the nature of the task, and they mediate different functions based on their inputs and outputs.32021 Although functions are compartmentalized within the cerebellum, the function of each cerebellar lobule is not well understood.
Structural anomalies and agenesis of the posterior vermis are related to cognitive deficiency and emotional symptoms.2223 The volume of vermian lobules VI and VII (also called the declive, folium, and tuber) is significantly correlated with the performance in cognitive testing.24 Vermian lobule VI is related to retrieval in the Sternberg working memory task,25 and vermian lobule VII is involved in the affective features of cerebellar cognitive affective syndrome and posterior fossa syndrome.2627 In addition, vermian lobules VI and VII have been found to be significantly smaller in patients with autism.28 These lobules might play roles in the regulation of cognition and emotion. There is evidence from recent neuroimaging and clinical studies that these cerebellar regions are critically associated with balance control and saccadic adaptation.
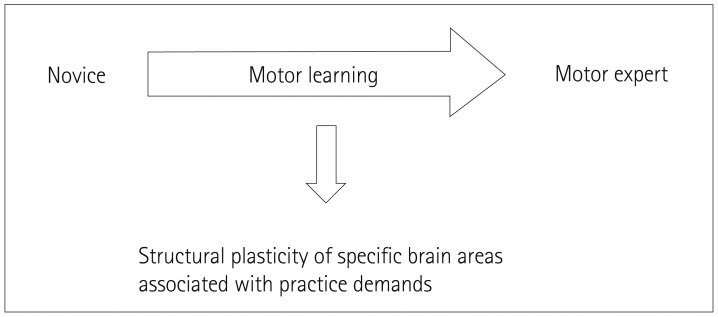
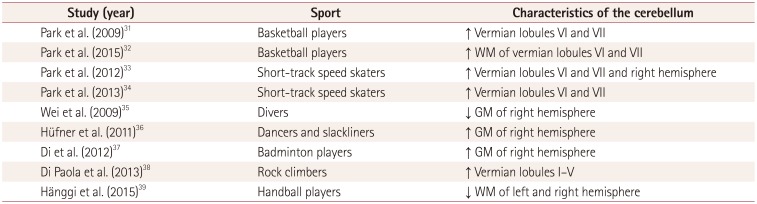
The function of the human cerebellum was traditionally elucidated using lesion studies. Cerebellar lesions induce specific behavioral impairments. Conversely, experience-dependent structural plasticity is helpful for providing insights into structure-function relationships in the human brain (Fig. 1). Longitudinal studies of juggling training demonstrated that motor learning induces structural changes in brain regions demanded by the task and suggest a causal relationship between motor learning and morphological plasticity.2930 Imaging studies have revealed structural alterations in the cerebellum of sportspeople after they have undergone intense training of complex motor skills (Table 1).313233343536373839 Such sports related structural changes in the cerebellum of sportspeople provide useful information for comprehending the specific function of each cerebellar lobule as well as neural mechanisms in motor experts. This review focuses on the function of vermian lobules VI and VII associated with balance control and eye movements, including in sportspeople.
BALANCE CONTROL
Lesion studies of both animals and humans suggest that the cerebellum, especially the medial zone (vermis and NF), plays a vital role in balance control.1112404142434445 The medial zone has inputs from the primary vestibular afferents and vestibular nuclei, reticular nuclei, pontine nuclei, and the spinal cord.46474849 The medial zone projects mainly to vestibular and reticular nuclei through the NF.4950 Animal studies have shown that the medial zone is essential for maintaining balance and postural tone and for walking.1140414551 In contrast, lesions in the lateral cerebellum (including the DN) cause no balance anomalies, very mild gait defects, but severe defects in the control of voluntary limb movements.1152 The lateral zone has inputs mainly from cerebral cortices with the thickest projections from the primary motor, premotor, primary somatosensory, posterior parietal, prefrontal cortices, and temporal lobe regions.4953 The lateral zone projects to the red nucleus through the DN and to cerebral cortices including primary motor, premotor, parietal, and prefrontal cortices through the thalamus.49505455 Recent track tracing investigation in non-human primates found that the primary motor cortex and several cortical motor areas (the supplementary motor, dorsal cingulate motor, and the ventral cingulate motor areas) project to lobules V–VIII of the vermis.56 This suggests that these regions of the vermis are where the cortical motor areas affect the descending control systems associated with the control of whole-body posture.
A human study using magnetic resonance imaging (MRI) revealed that lesions containing the NF and neighboring NI are involved in balance-control deficits.424357 Lesions in the NF that also include the NI are correlated with posture ataxia and gait deficits.58 Furthermore, the volume of the medial and neighboring intermediate cerebellum is negatively correlated with posture and gait deficits.59 Previous animal studies showed that the NF is involved in responses to vestibular stimulation6061 and controls limb extensor muscle tone for maintaining posture.4162 This suggests that the cerebellar vermis and fastigial nuclei play an essential role in balance control.
MRI-based lesion symptom mapping has also revealed that lesions involving the NF, anterior vermal lobules I–III, and posterior vermal lobules VIII and IX are associated with disorders of balance control.424357 Lesions of the vermal and paravermal lobules II, III, and IV are correlated with posture and gait ataxia.58 Pathology of the anterior superior cerebellar vermis induces balance and gait deficits in detoxified alcoholics.636465 In addition, the sway path length and sway prominence in the 2–5 Hz band are associated with the volume of the anterior-superior vermis in alcoholics.66
It is thought that tasks requiring sophisticated control of balance, such as tandem stance and standing on a cushion, are associated with the posterior lobe of the cerebellar vermis, whereas pathology in the anterior lobe of the cerebellar vermis is involved in balance deficits. Parallel fibers cross the midline cerebellar cortex and attach to Purkinje cells on either side of the posterior vermis. Purkinje cells might be critical for bilateral coordination of the legs and trunk, which is an essential function in balance control.45 In addition, the posterior vermis gets sparse peripheral somatosensory inputs, vestibular inputs, and some corticopontine inputs from the visual areas, whereas the anterior vermis takes more somatosensory input and little vestibular input.6768 Because the posterior vermis takes principally vestibular information, it has been thought to control balance and gait.
Deficits in tandem stance with the eyes open and standing on a cushion with the eyes closed are associated with lesions in vermal lobules VII and VIII, including the NF and NI.44 Five children with posterior inferior vermal splits ranging from lobules VI to X showed deficits in tandem gait and only slight anomalies in self-paced gait and hopping on one leg.12 Meanwhile, lesions involving vermian lobules VII, IX, and X cause lateropulsion.69
It is plausible that maintaining postural balance during unipedal gliding on smooth ice involves a more precise coordinating response to postural modification for shifting the center of gravity as in tandem stance than normal bipedal standing.7071 Recent structural MRI studies involving elite short-track speed skaters, who need exceptional balance control, indicated that their vermian lobules VI and VII are larger than in control subjects,3334 and that the volume of this region is correlated with the static balancing ability of the left leg in female skaters.34 This suggests that vermian lobules VI and VII play an essential role in controlling postural balance and reflect that this brain region is essential to balance function.
MOTOR LEARNING
Saccadic eye movements form an excellent motor learning model of the cerebellum because it is possible to slowly alter the amplitude of a saccade if the target moves constantly while the saccade is being performed.727374 Physiological and lesion researches involving non-human primates as well as imaging and lesion researches involving humans have revealed that the vermis especially vermian lobules VI and VII (declive, folium, and tuber) is a central area controlling saccadic adaptation. Alterations in the spike activities of Purkinje cells in vermian lobules VI and VII are correlated with the level of saccadic adaptation in primates.74 The saccadic area within vermian lobules VI and VII has been defined as the oculomotor vermis.7576
There is very strong evidence that vermian lobules VI and VII participate in the control of eye movements. The Purkinje cells of these lobules project to the caudal part of the NF, which in turn projects to the vestibular nuclei and saccade-related brainstem nuclei;77 indeed, this cerebellar region contains saccade-related neurons.78 Stimulation of this region of the vermis can evoke saccadic eye movements in both monkeys767980 and humans,81 whereas lesions in this region result in a permanent inability to adjust the amplitude of saccades.8283 Numerous studies performed over the past decade that have employed functional MRI (fMRI) and positron emission tomography have revealed the cerebellum to be associated with the control of eye movements. In humans, visually guided saccades activate vermian lobules VI and VII,848586 and saccadic adaptation is correlated with increased blood flow in these lobules.8788
Transiently impairment of the function of the posterior cerebellum (including vermian lobules VI and VII) using repetitive transcranial magnetic stimulation in healthy human subjects significantly weakened the ability to adjust the amplitude of saccades.89 Clinical studies have revealed that patients with focal cerebellar lesions or cerebellar degeneration are less able to adjust saccades.909192 In addition, lesions involving vermian lobules VII, IX, and X cause nystagmus.69
Motor learning probably underlies coordination between the eyes and hands and bimanual coordination.9394 Therefore, interactions between these two systems can be controlled exactly. An fMRI study demonstrated bimanual coordination-related activity within vermian lobule VI, suggesting its involvement in the executive function of bimanual coordination and the control of spatiotemporal complexity of coordination patterns.95 Coordination between eye and hand movements involves activation of vermian lobule VII.93 In addition, a three-dimensional MRI volumetric study proposed that greater volume of vermian lobules VI and VII in basketball players than in control subjects might be involved in the coordination between the eyes and hands and bimanual coordination, which are required for shooting and dribbling the ball.31
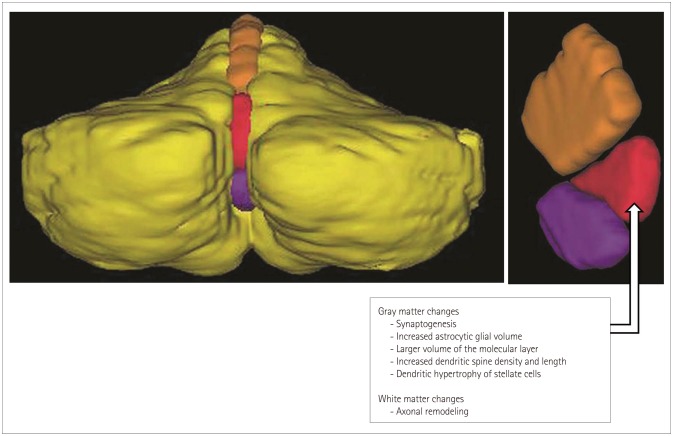
Several studies have revealed that learning acrobatic tasks that require substantial coordination of gross and fine motor skills not merely simple motor activity such as in locomotion or running drives synaptogenesis in the cerebellar cortex between the parallel fibers and dendritic spines of Purkinje cells.96979899100 In addition, increases in the astrocytic glial volume per Purkinje cell,101102 the volume of the molecular layer per Purkinje cell,96 and the dendritic spine density and length of Purkinje cells,103 as well as dendritic hypertrophy of stellate cells104 in the cerebellum have been observed after complex motor skill learning. Alterations in white matter involve the number and diameter of axons, myelin thickness, axon branching, axon trajectories, and myelination, and they effect the speed of impulse transmission.105 These might underlie the increased volume of vermian lobules VI and VII in sportspeople at the structural level (Fig. 2). Furthermore, this lobule might be closely associated with the acquisition of exceptional, specialized skills under broad activation of other motor movements.
CONCLUSIONS
Both human and animal studies indicate that vermian lobules VI and VII (declive, folium, and tuber) of the cerebellum might be associated with various features of balance control and motor learning, such as visually guided saccades and motor coordination, including coordination between the eyes and hands and bimanual coordination. Understanding the functional roles of vermian lobules VI and VII may lead to insights into the anatomical foundations and clinical signs. The findings of the present review may help understand neuroimaging findings in the context of cerebellar contribution in a wide range of motor functions.
 XML Download
XML Download