

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Colorectal cancer (CRC) is one of the most frequently diagnosed cancer types, and it accounts for a high annual mortality rate worldwide [1]. Although the relative 5-year survival rate of CRC has increased globally, it is only about 65%–70%. Until now, pathological staging has been the most accurate predictor of CRC prognosis. Therefore, novel prognostic markers for CRC are needed to allow for accurate predictions. Runt-related transcription factors (RUNXs) are a family of proteins that are important in the normal development and differentiation of cells. In mammals, the RUNX family consists of 3 gene groups, namely RUNX1, RUNX2, and RUNX3 [2345]. The gene RUNX1 is located on human chromosome 21q22.12, and it is involved in hematopoiesis and the growth of megakaryocytes, T cells, and B cells. RUNX2 is located on chromosome 6p21; it is involved in bone differentiation and may thus be associated with bonerelated diseases [6]. Finally, RUNX3 is located in 1p36 and is thought to be a tumor suppressor gene [6]. Recent studies have shown that it is associated with various malignancies, such as stomach, liver, lung, and esophageal cancers [789101112131415].
Aberrant signaling through this pathway is thought to promote tumor development in several tissues. Typically, inactivation of RUNX3 originates from allele loss or gene-silencing due to hypermethylation in CpG islands of the promoter region. However, the clinical significance of RUNX3 inactivation remains unclear. In this study, we used tissue microarrays containing primary CRC tissues to investigate the hypermethylation and expression of RUNX3 in relation to tumor features and to patient outcomes.
METHODS
CRC tissue samples
In the current study, we used 62 CRC tissue samples and 10 normal mucosal tissue samples collected from 62 patients who were diagnosed with CRC and underwent surgery at Soonchunhyang University Hospital. CRC and normal mucosal tissue samples were taken from the specimens, and each sample was divided each into 2 parts. One part of the sample was placed in liquid nitrogen immediately, while the other was embedded in a paraffin block after fixation in cold 10% neutral buffered formalin for 24 hours. For the diagnosis of cancer, a 4-µm paraffin section was stained using hematoxylin-eosin dye, and the histopathological findings were interpreted by a pathologist.
DNA extraction and bisulfite modification
To perform methylation-specific polymerase chain reaction (MS-PCR), a G-spin Genomic DNA Extraction Kit (Intron, Seongnam, Korea) was used to extract DNA from the tissue samples according to the manufacturer's instructions. Approximately 300 ng of DNA was diluted in 30 µL of distilled water, to which 3 µL of 3 M NaOH was added. The mixture was incubated at 75℃ for 20 minutes and subsequently cooled on ice. The DNA was then treated with 20 µL of 10 mM hydroquinone (Sigma-Aldrich, St. Louis, MO, USA) and 500 µL of 4.8 M sodium bisulfite (Sigma-Aldrich) and stirred well. This was followed by a 5-hour incubation at 55℃. Modified DNA impurities were removed using a Wizard DNA Clean-Up Kit (Promega, Madison, WI, USA). The mixture was then stored at room temperature using 0.3 M NaOH. Finally, DNA was precipitated using 100% ethanol and subsequently dried. Dried DNA was melted in 20 µL of distilled water prior to use as a template in PCR.
Methylation-specific polymerase chain reaction
Hypermethylation of the RUNX3 promoter was investigated via MS-PCR using methylation-specific primers. The primer sequences used to examine the methylated RUNX3 promoter were 5′-TTA CGA GGG GCG GTC GTA CGC GGG-3′ (sense) and 5′-AAA ACG ACC GAC GCG AAC GCC TCC-3′ (antisense), while those used to examine the unmethylated RUNX3 promoter were 5′-TTA TGA GGG GTG GTT GTA TGT GGG-3′ (sense) and 5′- AAA ACA ACC AAC ACA AAC ACC TCC-3′ (antisense). A series of reactions were performed in 35 cycles using the following PCR conditions: 2 minutes at 50℃, heat denaturation reaction for 10 minutes at 95℃, denaturation for 15 seconds at 95℃, annealing for 15 seconds at 55℃, and extension for 30 seconds at 72℃. The PCR products were confirmed using 10% TBE acrylamide gel. Positive controls for these reactions utilized universally methylated DNA (InterGen, Burlington, MA, USA). Distilled water was used instead of the DNA template in the negative control.
RNA extraction and reverse transcriptase-PCR
Total RNA was extracted from tissue samples using the TRizol Reagent (Life Technologies Inc., Grand Island, NY, USA) according to the manufacturer's instruction. Total extracted mRNA was purified using the RNeasy Mini kit (Qiagen, Hilden, Germany) and treated by RNase-free DNA I to remove genomic DNA contamination. Five hundred ng/µL of mRNA was reverse transcribed using a ReverTraAce qPCR RT kit (Toyobo, Tokyo, Japan) and reverse transcription PCR was performed using the Maxim PCR PreMix i-starTaq kit (Intron) according to the manufacturer's instruction. The following primers were used for reverse transcriptase (RT)-PCR: human RUNX3 sense; 5′-GAG TTT CAC CCT GAC CAT CAC T GT G-3′, antisense; 5′-GCC CAT CAC TGG TCT TGA AGG TTG T-3′ and human GAPDH sense; 5′-ACG ACC ACT TTG TCA AGC TC-3′, antisense; 5′-TCT TCC TCT TGT GCT CTT GC-3′. PCR products were analyzed and visualized by 2% agarose gel electrophoresis.
Immunohistochemical staining
To examine the correlation between hypermethylation of the RUNX3 promoter and RUNX3 protein expression, immunohistochemical (IHC) staining was performed. To retrieve antigen, sectioned samples on slides were placed in 0.01 M sodium citrate buffer (pH, 6.0) and boiled in a microwave for 20 minutes. Samples were then cooled to room temperature for more than 1 hour and rinsed three times (3 minutes each) in distilled water. To inhibit the intrinsic activity of peroxidase, methanol was used to prepare a 3% hydrogen peroxide solution, and the tissue samples were incubated in this solution at room temperature for 30 minutes. The tissue samples were then rinsed with distilled water 3 times, following which they were washed 3 times with phosphate-buffered saline (PBS; pH, 7.4). The samples were then incubated with the primary antibody (1:100) for 1 hour at room temperature. Again, the sample was rinsed with PBS three times (3 minutes each). The tissue samples were incubated with a PicTure-Plus Kit (Zymed Laboratories Inc., South San Francisco, CA, USA) for 30 minutes at room temperature. After the samples were rinsed with PBS three times, diaminobenzidine was added to the samples; following this, they were counterstained with hematoxylin dye and examined using light microscopy.
IHC analysis
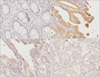
RUNX3 stained tissue cores were examined by 2 independent pathologists, and a consensus score was determined for each specimen. A positive reaction the for antibody was scored into four categories, as follows: 0 (0%), 1+ (1%–33%), 2+ (34%–66%), and 3+ (67%–100%). RUNX3 expression was defined to be expressed in the nucleus or cytoplasm if the intensity was estimated at ≥2 (Fig. 1).
Statistical analysis
The correlations between hypermethylation/expression of RUNX3 and clinicopathologic parameters were evaluated using the chi-square or Fisher exact test. The overall survival was calculated as the first diagnosis of CRC to death from any other cause or until April 2016. Survival curves were estimated using the Kaplan-Meier method. All P-values quoted were 2-sided, and P < 0.05 was considered to indicate a statistically significant difference. All analyses were performed using SPSS ver. 17.0 (SPSS Inc., Chicago, IL, USA).
RESULTS
Patient characteristics
Between January 2008 and February 2009, 62 patients with a median age of 64 years (range, 31–83 years) were included in the study. There were 37 men (59.7%) and 25 women (40.3%). Two patients (3.2%) were in stage IV. The most common histological grade of tumor was moderate differentiation (51 patients, 82.3%). The baseline characteristics are listed Table 1.
Prevalence of RUNX3 hypermethylation
MS-PCR was performed in 62 CRC tissue samples and 10 normal colon tissue samples (Fig. 2). Of the 62 CRC tissue samples, 20 (32.3%) presented hypermethylated RUNX3 promoters, while the remaining 42 (67.7%) showed no methylation. All 10 normal colon tissue samples showed no methylation.
Relationship between hypermethylation and mRNA expression of RUNX3
To examine the correlation between hypermethylation and mRNA expression of RUNX3, RT-PCR was performed for all CRC tissue samples, as well as the 10 normal colon tissue samples (Fig. 3). The mRNA expression of RUNX3 was observed in all 10 normal colon tissue samples. However, of the 20 samples that showed RUNX3 promoter hypermethylation, 14 (70.0%) exhibited decreased expression of RUNX3 compared to normal colorectal tissue samples, while the remaining 6 samples (30.0%) showed normal RUNX3 expression. In contrast, of the 42 CRC tissue samples that showed no RUNX3 promoter hypermethylation, 27 (64.3%) demonstrated normal mRNA expression and 15 (35.7%) exhibited decreased expression. Hypermethylation of CpG islands of the promoter region of RUNX3 was significantly associated with expression of RUNX3 mRNA (P = 0.011) (Table 1).
Association between promoter hypermethylation of RUNX3 and clinicopathological parameters
When the tumors of the 62 patients were divided according to pathological stage classification, stage was not associated with RUNX3 methylation status (P = 0.307). We also found that differentiation of cancer was not correlated with hypermethylated RUNX3 (P = 0.179). However, promoter hypermethylation of RUNX3 was significantly associated with vascular invasion (P = 0.006) and lymphatic invasion (P = 0.002) (Table 1). In contrast, other clinicopathological variables showed no correlation with hypermethylated RUNX3.
Hypermethylation/expression of RUNX3 and overall survival
The median follow-up duration of the 62 CRC patients was 81.3 months (range, 8.9–96 months). Eighteen patients died during the follow-up period. All patients died from cancer progression. Compared to unmethylation, hypermethylation of RUNX3 was significantly associated with worse CRC prognosis (P = 0.038, log-rank test) (Fig. 4). Promotor hypermethylation was significantly associated with expression of RUNX3 (P = 0.019). However, RUNX3 expression in patients was not associated with CRC prognosis (P = 0.363, log-rank test) (Fig. 5).
DISCUSSION
The molecular mechanisms underlying the malignant transformation of colorectal tissues can be divided into the typical adenoma-carcinoma sequence and the acquired genetic changes due to promoter hypermethylation of tumor suppressor genes [16]. The genes that are associated with the acquired genetic changes include p16 and Rb, which are involved in controlling the cell cycle; E-cadherin and TIMP-3, which are involved in cell attachment; and hMLH1, BRCA1, MGMT, and p14, which are involved in DNA repair. These genes act as etiological factors for cancer when their expression is decreased by the hypermethylation of promoters [17]. The RUNX3 gene encodes a tumor suppressor protein and is located on chromosome 1p36. It plays a role as a regulator of mRNA transcription along with p300 in the tumor growth factor β (TGF-β) signaling pathway [18]. TGF-β is involved in functions like pattern formation, cell behavior, differentiation, proliferation, and apoptosis during fetal development. In addition, it is involved in repairing tissue injury and regulating immune cells in adults [18]. TGF-β binds to type II receptors and type I receptors within the cytoplasm in a sequential manner, after which it becomes activated [18]. Once activated, it phosphorylates Smad proteins [19]. Phosphorylated Smad binds to Smad4, which then translocates to the nucleus. This complex then binds to the promoter region of a target gene, where it induces expression. Currently, transcription factors like p300/CBP transcriptional coactivator or RUNX are synchronously involved. For the activation of tumor suppressor genes in the TGF-β signaling pathway, the tumor-suppressing functions of the complex composed of Smad2, Smad4, p300, and RUNX3 must be well harmonized [20].
In stomach cancer tissue, the inactivation of RUNX3 has been reported to occur because of the hypermethylation of CpG islands within the promoter region. A correlation between the malignant transformation of stomach tissue and RUNX3 expression was identified in 8.1% of patients with gastritis, 27.3% of those with gastric adenoma, and 64.0% of those with stomach cancer [9]. These results suggest that malignant transformation of many cell types may be associated with promoter methylation status [9]. In the current study, MSPCR was performed for the promoter region of RUNX3, which revealed promoter hypermethylation in 32.3% of CRC tissue samples. In normal colon tissue samples, no methylation was detected. These proportions are lower than those reported by Oshimo et al. [8], who observed hypermethylation in 71.0% of stomach cancer tissue samples. In recent years, several follow-up studies have been conducted to examine the hypermethylation of RUNX3 promoter CpG islands.
The correlation between the status of RUNX3 promoter methylation and the expression of mRNA was also assessed, which showed that mRNA was not expressed in 14 of 20 CRC tissue samples (70.0%) with hypermethylated promoter regions, but it was detected in the remaining 6 samples (30.0%). Of the 42 samples that showed no methylation of the RUNX3 promoter, 27 (64.3%) exhibited expression of RUNX3 mRNA, while 15 (35.7%) did not (Table 1). Comparing these results to those of prior publications, a smaller proportion of cancer cells showed no mRNA expression. Tumor heterogeneity may be responsible for these findings.
Little is known about the correlations between the clinical characteristics of CRC and hypermethylation of the RUNX3 promoter. Some studies have described the above correlation in patients with stomach cancer or urinary bladder cancer. However, in patients with hepatoma, no statistically significant association has been reported [1221]. Recent studies have determined that hypermethylation or expression of RUNX3 is highly associated with higher pathological staging and poor prognosis in various tumors, including CRC. However, these results have not been consistent. In addition, only 2 studies estimated the relationship between overall survival and RUNX3 hypermethylation/expression in CRC [22]. Ogino et al. [23] reported that CRC patients with RUNX3 hypermethylation had a poorer 1-year survival than those without RUNX3 hypermethylation (P < 0.001). However, the follow-up duration was only 1 year. In this study, we found that hypermethylation of RUNX3 was significantly associated with worse prognosis of CRC after 8 years of follow-up (P = 0.038).
Contrary to our expectation, the present data showed that RUNX3 expression was not associated with CRC prognosis (P = 0.363). Soong et al. [24] observed that RUNX3 expression was associated with better survival than no RUNX3 expression (P = 0.025). Interestingly, in this study, cytoplasmic expression of RUNX3 (only 9% of the study) was associated with worse prognosis. However, patients with no expression of RUNX3 showed similar prognoses to those with RUNX3 nuclear expression. To accurately determine the association of prognosis with RUNX3 expression, we need to analyse the RUNX3 expression in the nucleus or cytoplasm in further studies. The associations between RUNX3 hypermethylation and the clinicopathologic features of CRC are shown in Table 1. No significant association was seen with age, sex, histological grade, or stage. However, hypermethylation of RUNX3 was found to be associated with vascular (P = 0.006) and lymphatic (P = 0.002) invasion.
The limitations of the present study include its retrospective design and relatively small sample size. It could have been affected by potential selection bias. In conclusion, the major finding of this study was that hypermethylation of RUNX3 may be a predictor of a poor prognosis in CRC cases. However, further large-scale studies are warranted to determine the clinical significance, including prognosis, of RUNX3 hypermethylation/ expression in CRC.





 XML Download
XML Download