

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Among alternative laboratory models, pigs are most suitable because they have cardiovascular, renal, and gastrointestinal organs similar to those of humans [2136]. However, transgenic (TG) pig production efficiency is very low compared to that of other mammals such as cattle, sheep, and mice due to incomplete reprogramming of somatic cell nuclear transfer (SCNT) embryos [32]. Therefore, unstable development and unsuccessful implantation of SCNT embryos limit the production of TG pigs [2338]. In genetically engineered animals, early development of embryos has been reported to be affected by abnormal expression of foreign genes, causing miscarriage or early mortality of TG animals [35]. To overcome this limitation, many researchers have recently applied conditional transgene expression systems such as the ecdysone-inducible gene switch, tetracycline-inducible system, or Gal4/UAS system to prevent inhibition of embryonic development and to induce gene expression as desired by the researcher [2033]. These systems have begun to be used in large animals, such as pigs, in addition to their original use in rodents. In 2006, the first TG pigs with a tetracycline-inducible system were produced by a pronuclear DNA microinjection technique [25].
The CreER-LoxP system is a frequently used site-specific recombination system for averting undesirable development outcomes such as embryonic or postnatal lethality [2631]. Cre recombinase recognizes two consensus sequences, called locus of X(cross)-over in P1(LoxP), and catalyzes excision, insertion, inversion, or even translocation of target DNA sequences in accordance with their location and orientation [12]. In addition, CreERT2, a fusion protein of Cre recombinase and the modified estrogen receptor (ER) with a higher sensitivity to its optimal ligand 4-hydroxytamoxifen (TM) than to endogenous estradiol, was developed to control the timing of a recombination event by treatment with the ligand [1030]. This system has been widely applied to TG mouse models that require conditional activation or knock-out of genes of interest [39].
Glial fibrillary acidic protein (GFAP), a class III intermediate filament protein, is an astrocytic cell lineage marker [7]. A 2.2 kb 5′-flanking sequence of the human GFAP (hGFAP) gene, called Gfa2, has been identified and used to induce the expression of oncogenes specifically in astrocytes in numerous brain tumor mouse models [217]. However, the mouse brain is 750 times smaller than the human brain, and it includes a much lower amount of white matter than that in the human brain. Furthermore, the mouse brain is lissencephalic (i.e., it has a smooth outer cerebral cortex). In contrast, the pig brain is 7.5 times smaller than the human brain, has a white-to-gray matter ratio (60:40) similar to that of humans, and is gyrencephalic (i.e., it has a highly convoluted outer cerebral cortex), similar to the human brain. Thus, the pig is a better animal model than the mouse for research in human brain diseases [1416].
In the present study, we constructed pig GFAP promoter-CreERT2 [8] and enhanced green fluorescent protein (EGFP)-LoxP transgenes for use in TM-inducible CreERT2-mediated recombination. We then established donor TG pig fibroblasts with pGFAP-CreERT2/LCMV-EGFPLoxP transgenes for use in SCNT. After SCNT, we evaluated the effect of the transgenes on TG-SCNT embryo development. After embryo transfer, we generated 5 TG pigs and confirmed the presence of the transgenes and TM-inducible CreERT2-mediated recombination in a living TG pig.
Materials and Methods
Ethics statement
The experimental protocol was approved by the Committee on Ethics of Animal Experiments of the Chungbuk National University (permission No. CBNUR-871-15). All surgeries were performed under anesthesia, and we tried to minimize the suffering of the animals.
Chemicals
The equipment and reagents used in this study are detailed in the following sections. All other chemicals were purchased from Sigma-Aldrich (USA).
Vector construction
To construct pCDH-pGFAP-MCS-EF1-puro, the CMV promoter sequence of the pCDH-CMV-MCS-EF1-puro lentiviral vector (System Biosciences, USA) was removed by using two restriction enzymes, SnaBI and NheI. The pGFAP promoter sequence (−1,817 to −17 upstream from the pGFAP transcription start site +1) in the pGL3-basic-pGFAP promoter plasmid [8] was digested with EcoRI (blunted by the Klenow fragment) and NheI. Subsequently, the pGFAP promoter sequence was ligated into the aforementioned CMV promoter-deleted lentiviral vector. Finally, to construct a lentiviral vector containing the pGFAP-CreERT2 gene construct, the CreERT2 gene digested by EcoRI from the pCre-ERT2 plasmid [19] was inserted into the identical restriction enzyme sites of pCDH-pGFAP-MCS-EF1-puro.
To generate the CMV-LoxP-EGFP-pA-LoxP construct, LCMV:EGFP(LoxP)MCS (No. 31377; Addgene, USA) was modified by removing the EGFP sequence by using NcoI and BsrGI. Then, the EGFP sequence from EGFP-C2 (No. 6083-1; Clontech Laboratories, USA) that had been digested with the same restriction enzymes was inserted into the aforementioned EGFP-deleted vector.
Lentivirus production
To produce the lentivirus, 293FT cells were plated on 100-mm plates in complete Dulbecco's modified Eagle medium (DMEM) with 10% fetal bovine serum (FBS; Gibco, Australia) and 2 mM L-glutamine (Lonza, Switzerland) and allowed to adhere for 12 h. Transfections were performed as per the manufacturer's instructions with 4 µg of transfer vector plasmid, 6 µg of third generation packaging plasmids, and 24 µL Polyexpress agent (Excellgene, USA) in serum-free DMEM (Invitrogen, USA). Six hours after transfection, the cells were washed with phosphate-buffered saline, after which, fresh complete DMEM with L-glutamine and 10% FBS was added, and the cells were incubated for an additional 42 h. Supernatant medium containing lentivirus was collected 48 h after transfection and filtered through a 0.45-µm syringe. Each lentivirus was concentrated with the Lenti-X Concentrator (Clontech Laboratories) according to the manufacturer's instructions.
Donor cell line construction
The primary cell line was derived from male Yucatan miniature pig fetal fibroblast cells. The cell line was named Cloud male. It was transfected with LCMV-EGFPLoxP using an electroporation tool (Neon Transfection System; Invitrogen) according to the manufacturer's instructions. After 3 to 4 weeks, EGFP-positive cells were sorted by flow cytometry (fluorescence-activated cell sorter [FACS], Aria II; BD Biosciences, USA). The EGFP-positive cells were then infected with the pCDH-pGFAP-CreERT2-EF1-puro lentivirus vector by using polybrene (6 µg/mL). Three days after infection, cells were selected by puromycin treatment (2 µg/mL; Clontech Laboratories) for 5 days.
Oocyte collection and in vitro maturation
Oocyte collection and in vitro maturation was performed according to Hwang et al. [18]. Briefly, porcine ovaries were collected from a slaughterhouse. Porcine follicular fluid (pFF) and cumulus-oocyte complexes (COCs) were recovered from 3 to 6 mm ovarian follicles by aspiration. The composition of the medium used during in vitro maturation (IVM) was as follows: TCM199 (Gibco), 0.6 mM cysteine, 0.91 mM sodium pyruvate, 10 ng/mL EGF, 75 µg/mL kanamycin, 1 µg/mL insulin, and 10% (v/v) pFF. Porcine COCs were co-cultured at 50 to 60 cells per well in a 4-well dish (Nunc, Denmark) with 500 µL IVM media. The conditions for IVM were 39℃ in a 5% CO2 atmosphere in a humid incubator (Astec, Japan). Maturation was performed in IVM medium with 10 IU/mL equine chorionic gonadotropin and 10 IU/mL human CG for 22 h. The cells were then moved into hormone-free IVM medium and cultured for 18 h. Matured COCs were denuded by using gentle pipetting with 0.1% hyaluronidase and HEPES-buffered Tyrode's medium containing 0.05% (w/v) polyvinyl alcohol (TLH-PVA) medium. Denuded oocytes obtained through this process were used in subsequent experiments.
SCNT and in vitro culture
After 40 h of IVM, denuded oocytes at metaphase II (MII) stage were chosen for enucleation. MII oocytes were washed thrice in calcium-free TLH containing 0.2% bovine serum albumin (TLH-BSA) and 5 µg/mL cytochalasin B (CB). Enucleation was performed by using a micro-manipulator with a 16-mm glass pipette (Humagen, USA). After enucleation, trypsinized TG donor cells were transferred into the perivitelline space of enucleated oocytes. Next, they were fused by two pulses of 180 V/mm direct current for 60 µsec in a 260 mM mannitol solution containing 0.1 mM CaCl2 and 0.05 mM MgCl2 by using a cell fusion generator (LF201; Nepa Gene, Japan). After electrical fusion, the SCNT embryos were incubated in 6-dimethyl aminopurine with 5 µg/mL CB in 30-µL droplets of porcine zygote medium (PZM) for 4 h (post-activation). Embryos were transferred to PZM droplets for in vitro culture. On the second day after fusion, embryo cleavage was evaluated (1 cell, 2-3 cell, 4-5 cell, 6-8 cell stages and fragmented embryos), and embryos were transferred to new PZM droplets. On the 4th day, we transferred the embryos to PZM droplets containing 10% FBS. On the 7th day after fusion, blastocyst (BL) formation was evaluated quantitatively (early, expanded, hatched BL).
Embryo transfer and induction of CreERT2 system
Non-superovulated, naturally cycling Landrace × Duroc crossbreed gilts were used as surrogate mothers. The ear veins were injected with 1 mL ketamine (50 mg/mL; Yuhan, Korea) and 3 mL xylazine (100 mg/mL; SF, Korea) for preanesthesia. Thereafter, respiratory anesthesia using isoflurane liquid (Hana Pharm, Korea) was maintained. All surgical instruments were sterilized before surgery. A midventral laparotomy was performed for exposure of the reproductive organs. At 4 h post-activation, SCNT embryos (100–150 embryos per recipient) were transferred into surrogate oviducts (at the ampullary-isthmic junction). After 30 days, ultrasonography was used to diagnose pregnancy.
In previous studies, CreERT2-mediated recombination in mice was induced by oral or intraperitoneal injection. However, injections by oral or intraperitoneal injection. Compared with oral administration, intraperitoneal injections require a 5 to 10 fold increase in concentration [11]. Therefore, we decided to induce CreERT2-mediated recombination in piglets by oral administration. Previously, the tamoxifen concentration in blood after oral administration of tamoxifen was reported in rats [22]. We established the concentration on the basis of this result. At 3 months of age, the piglet was orally administered 15 mg/kg TM for 5 days and then euthanized 7 days later.
Genomic DNA isolation and polymerase chain reaction analysis
Genomic DNA was isolated from frozen tissues and cells by using the Wizard genomic DNA purification kit (Promega, USA) according to the manufacturer's instructions. Large and tough tissues were homogenized by using a Bullet Blender (Next Advance, USA) according to the manufacturer's instructions. Transgenes in genomic DNA were amplified with the appropriate primer set (EGFP, CreERT2, or 18S rRNA) and NeoTherm DNA polymerase (Genecraft, Germany) under the following thermal cycling conditions: 60 sec at 95℃; 30 cycles of 30 sec at 95℃, 40 sec at 58℃, and 60 sec at 72℃; followed by 10 min at 72℃.
For semi-nested polymerase chain reaction (PCR) analysis to detect CreERT2-mediated recombination, first-round PCR was performed by using the first pair of primers (forward and reverse) and NeoTherm DNA polymerase under the following thermal cycling conditions: 60 sec at 95℃; 35 cycles of 30 sec at 95℃, 30 sec at 56℃, and 120 sec at 72℃; followed by 10 min at 72℃. All PCR products were separated by DNA electrophoresis in a 1% agarose gel. Gel blocks at the expected product size after recombination were removed without UV exposure. Then, in-gel DNA extraction in the isolated gel slices was performed by using a gel extraction kit (Elpis Biotech, Korea) according to the manufacturer's instructions. Finally, second-round PCR was performed by using forward and reverse primers under PCR conditions identical to those of the first-round PCR. The primer sequences are presented in Table 1.
Quantitative reverse-transcription PCR
Quantitative reverse-transcription PCR (qRT-PCR) was performed by using Takara SYBR Premix Ex Taq (Takara Bio, Japan) and CFX096 (Bio-Rad, USA) to determine mRNA levels. Total RNA was isolated from cells by using TRIzol reagent (Invitrogen) according to the manufacturer's instructions. RNase-free DNase-treated RNA (1 µg) was used as the template to synthesize cDNA by using the RevertAid first-strand cDNA synthesis kit (Thermo Fisher Scientific, USA) according to the manufacturer's instructions. The expression levels of genes of interest were normalized to that of 18S rRNA. All primer sequences are presented in Table 1.
Tissue immunofluorescence assay
Pig brain tissues embedded in paraffin were sliced into 4-µm-thick sections, deparaffinized, and rehydrated as follows: 3 changes of xylene (5 min each), 2 changes of 100% ethanol (3 min each), 95% ethanol (3 min), 80% ethanol (3 min), and 70% ethanol (3 min). Then, antigen retrieval was performed by using sodium citrate buffer (10 mM sodium citrate, 0.05% Tween-20, pH 6.0) followed by microwaving for 20 min. Pig brain tissues were stained with primary antibodies against pGFAP (691102, 1:500; MP Biomedical, USA), Cre recombinase (12830S, 1:200; Cell Signaling, USA), and EGFP (AB6673, 1:200 and AB290, 1:200; Abcam, UK) for 12 h at 4℃. This was followed by incubation with secondary antibodies conjugated with Alexa Fluor 488 or 594 (A11001, A11005, A11012, or A11055, all 1:500; Thermo Fisher Scientific) for 1 h at room temperature. Nuclei were then stained with 4′,6-diamidino-2-phenylindole (1 µg/mL) for 5 min. Fluorescence images were obtained by using a confocal laser scanning microscope (LSM 5 Pascal; Carl Zeiss Vision, Germany).
Statistical analysis
Experiments for statistical analysis were replicated more than three times. All data were analyzed by one-way ANOVA followed by Duncan's test as provided in IBM SPSS Statistics software (ver. 21; IBM, USA). Mean ± SEM values were obtained. Differences were considered significant at p < 0.05.
Results
Construction of donor TG pig fibroblasts with the astrocyte-specific pGFAP-CreERT2/EGFPLoxP recombination system
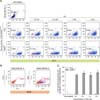
Before constructing a pig astrocyte-specific recombination system, we tested whether the CreERT2/loxP system worked in pig fibroblasts by detecting EGFP-floxed populations after TM-mediated CreERT2 activation (Fig. 1). Subsequently, we constructed an astrocyte-specific pGFAP-CreERT2/EGFPLoxP recombination vector for developing TG pigs. Using these vectors, we attempted to construct nuclear donor fibroblast cells with pGFAP-CreERT2/EGFPLoxP transgene constructs. Cells expressing the EGFP transgene were sorted by FACS, and EGFP expression was validated by fluorescence microscopy (Fig. 2). Next, we infected the pGFAP-CreERT2-puro lentiviral vector into cells expressing the EGFP transgene, and three cell colonies were established as final donor cells for SCNT by using antibiotic selection. The presence of each transgene into the pig fibroblast genome was identified by performing genomic DNA PCR analysis (panel D in Fig. 2). In the same manner, we constructed nuclear donor fibroblasts with an astrocyte-specific CreERT2/LoxP recombination system by using two vectors and confirmed the presence of each transgene of the system in the genomic DNA.
Development of SCNT embryos in vitro
The development of SCNT embryos was evaluated in order to assess the normality of the TG donor cell line (Fig. 3). Cloud male No. 5 (primary cell line from a normal fetal Yucatan minipig), and Cloud male No. 5 pGFAP-CreERT2 + LCMV-EGFPLoxP (vector-inserted cell line) cells were used as the donor cells. The fusion rates (71.8 ± 6.1 and 74.2 ± 2.6, respectively) and cleavage rates (73.8 ± 4.0 and 63.6 ± 2.2, respectively) did not show significant differences. The BL formation rate was not significantly different between the Cloud male No. 5 and Cloud male No. 5 pGFAP-CreERT2 + LCMV-EGFPLoxP cells (19.5 ± 1.2 and 21.0 ± 1.6, respectively). These findings indicate that the foreign genes in the donor cells did not affect the development of SCNT embryos.
Production of TG cloned pigs using Cloud male No. 5 pGFAP-CreERT2+LCMV-EGFPLoxP donor cells
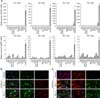
Embryo transfer was performed thrice just before the onset of ovulation of the surrogate mothers (panel A in Fig. 4). One of the surrogates became pregnant and delivered 5 TG piglets at 115 days after pregnancy. Three piglets (No. 1, 2, and 3) were stillborn, No. 5 died after 22 days, and only No. 4 survived. Genomic DNA analysis in skin cells confirmed that all 5 piglets had the pGFAP-CreERT2/EGFPLoxP transgene constructs (panels B and C in Fig. 4). Further diagnosis of stillbirths (No. 1, 2, and 3) was not performed. After 3 months, the surviving piglet was orally administered 15 mg/kg TM for 5 days and then euthanized 7 days later.
Expression of the pGFAP-CreERT2/EGFPLoxP transgene constructs in the nervous system of TG piglets
To determine whether the transgene controlled by the pGFAP promoter sequence was expressed specifically in astrocytic lineages, tissues from 4 TG piglets, including two spontaneously aborted TG piglets (No. 2 and 3), were analyzed by qRT-PCR. The carcass of TG pig No. 1 was unsuitable for the experiments because of inappropriate storage; therefore, it was excluded.
Endogenous GFAP expression was primarily restricted to brain tissues, and the CreERT2 gene was also highly expressed in brain tissues compared to that in other tissues (panel A in Fig. 5). Immunofluorescence (IF) staining was performed to determine whether CreERT2 controlled by the pGFAP promoter was expressed in GFAP-positive glial cells. CreERT2 expression was detected not only in star-shaped GFAP-positive cells but also in some other cell types (panel B in Fig. 5). To determine whether the pGFAP promoter transcribed the CreERT2 transgene in non-astrocytic cells, fibroblasts obtained from the ear tissues of aborted TG pigs No. 2 and 3 were assessed by primary culture. The cell lines were treated with TM. We observed that CreERT2 controlled by the pGFAP promoter was not expressed in these cells (Fig. 6). Furthermore, EGFP transgene expression in brain tissues was detected by IF staining (panel C in Fig. 5). Although GFAP-positive cells exhibited a slightly weaker fluorescence intensity than other cell types in brain tissue, most of the cells expressed EGFP. These data indicate that the pGFAP-CreERT2/EGFPLoxP developed in this study can be used as an astrocyte-specific recombination system.
pGFAP-CreERT2/EGFPLoxP recombination in GFAP-expressing cells after TM treatment
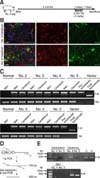
We next administered TM to TG pig No. 4 for 5 days and sacrificed it after one week to determine whether the pig astrocyte-specific pGFAP-CreERT2/EGFPLoxP recombination system was present (panel A in Fig. 7). We first used IF staining to compare the EGFP signal intensity in brain tissues of TM-treated TG pig No. 4 with that of untreated TG pig No. 5, which had died 22 days after birth and was used as a negative control for CreERT2-mediated recombination. Astrocytic cells without EGFP expression were mainly detected in the brain of pig No. 4 (panel B in Fig. 7).
We also conducted genomic DNA PCR analysis by amplifying the recombination site using forward and reverse primer pairs flanking the two LoxP sites. PCR products amplifying pre-recombinant DNA were only detected in brain and skin tissues (panel C in Fig. 7). We attributed the invisible post-recombinant PCR products in the PCR analysis to the presence of a very small population of recombined cells. Therefore, we used a semi-nested PCR analysis to increase PCR sensitivity. After first-round amplification, second-round PCR was performed with the first-round PCR product and a primer set that enabled amplification of an internal sequence of the first-round PCR product (panel D in Fig. 7). The presence of a 450-base pair PCR product indicated that post-recombination occurred only in the brain of the TM-treated pig (panel E in Fig. 7). These data indicate that pGFAP-CreERT2-driven recombination occurs in a very small population of cerebral astrocytic cells in response to TM treatment.
Discussion
The production and characterization process for pig models of human neurological disease is more expensive and time-consuming than that for mouse models [15]. The purpose of this study was to generate TG piglets using an astrocyte-specific pGFAP-CreERT2-inducible system and to confirm the effectiveness of the system in vivo.
TM-inducible recombination systems using the pGFAP promoter sequence have been widely used in astroglia-related studies requiring conditional turning on or off of transgenes. This is an important tool for the generation of various TG models, such as brain tumor models. In a previous study, we isolated a 5′-flanking sequence of pGFAP from the Yucatan miniature pig genome and verified its function as an astrocyte-specific promoter in vitro [8]. In the present study, we introduced an astrocyte-specific pGFAP-CreERT2 inducible system to prevent stillbirth and premature death caused by the expression of transgenes and the neural-specific pGFAP promoter. The hGFAP promoter has been widely used to produce mouse models for central nervous system (CNS) diseases [14].
Techniques for the production of TG pigs include SCNT by micromanipulation, handmade cloning (a simple variant of SCNT technology without micromanipulation), and microinjection, among others [63437]. Herein, we constructed an astrocyte-specific conditional pGFAP-CreERT2/EGFPLoxP recombination system and evaluated its validity in TG pig embryos and assessed the pig model generated by SCNT technology.
In this study, the production rate of TG pigs was approximately 1.9% (5 piglets/269 SCNT embryos). At present, the production efficiency of TG pigs is very low worldwide. When calculated based on birth weight, the efficiency is 1% to 2%; however, a production rate of 5.5% (28 piglets/511 SCNT embryos) was reported by Walker et al. [38]. There are several reasons for the low pregnancy and production rates. First, the current TG techniques used in the TG embryo creation process, such as IVM of oocytes and somatic cell treatment processing, are incomplete. Second, the low developmental potential of TG embryos and the physiological characteristics of pigs necessitate the implantation of at least 3 to 4 embryos [38]. Finally, transgenes have been reported to negatively affect embryonic and fetal development [3539]. Our present astrocyte-specific pGFAP-CreERT2 inducible system demonstrated no deleterious effect on the rate of embryo development in vitro. In this study, we produced SCNT embryos using a Cloud male No. 5 pGFAP-CreERT2 + LCMV-EGFPLoxP donor cell line that was verified in vitro. The embryos were transferred into a surrogate pig that gave birth to five pGFAP-CreERT2/EGFPLoxP TG piglets. In qRT-PCR results, we detected relatively high levels of GFAP and CreERT2 expression in the cerebellum relative to that in the cerebrum. This result might be attributed to the GFAP-expressing Bergmann glial cells, which are a major cell type in the cerebellum [5]. IF staining and semi-nested PCR analysis showed that the CreER-mediated astrocyte-specific recombination system was functional in some cerebral astrocytes after TM administration to TG pig No. 4. However, in this process, some technical limitations related to low recombination efficiency occurred. First, despite its cerebral-specific expression, CreERT2 regulated by the pGFAP promoter showed low expression compared to that of endogenous GFAP expression in the brain. Previously, in a TG mouse model, an intron sequence of mouse protamine-1 was added into the 3′ region of the Gfa2 transgene [2]. Therefore, in the TG pig model, it might also be necessary to introduce a transcription-enhancing element, such as an intron(s) or enhancer(s), to increase transgene expression. Second, EGFP transgene silencing was detected in TG pigs. The transcriptional activity of the CMV promoter becomes progressively weaker during long-term cell culture or the reprogramming process through promoter methylation. Kong et al. [24] reported that methylation of the CMV promoter caused a gradual decrease of GFP expression in TG pigs and pig fibroblasts. Therefore, the addition of stably expressing promoters such as elongation factor 1α or CMV early enhancer/chicken β-actin promoter should be considered [13]. Third, unstable expression and transgene silencing might result from positional effects of random integration [27]. The use of a homologous recombination-based locus-targeting knock-in system to integrate the transgene into a safe harbor locus, such as Rosa26, could be an effective alternative strategy [2829]. Finally, given that the turning off of the reporter expression in target cells was almost impossible to clearly distinguish and quantify, an effective dual fluorescence reporter system should be applied [28].
In summary, we showed the successful generation of a pGFAP promoter-driven, TM-inducible CreERT2-LoxP recombination system in a TG pig generated by SCNT. To our knowledge, this is the first report of an astrocyte-specific pGFAP-CreERT2 inducible system in TG pigs. Therefore, we suggest that this technology might be a useful tool for studying the physiology of astrocytes and generating TG pig models of CNS diseases such as Huntington's disease, Alzheimer's disease, and brain tumors.





 XML Download
XML Download