

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
The primary cilium is a microtubule-based solitary organelle that protrudes from the cell surface. It consists of a backbone of a microtubule, a 9+0 axoneme composed of α/β-tubulin heterodimers, which anchors to the basal body, and is surrounded by the plasma and ciliary membranes. The primary cilium senses diverse extracellular signals such as mechanical, photo, and biochemical signals and transduces them into the cell.123 Recent studies have demonstrated that the primary cilium plays a crucial role in the progression of various diseases in many organs, including the kidneys.456789
The primary cilium is typically 3 to 10 µm long in quiescent and fully differentiated cells.5691011 Its length is dynamically altered during the cell cycle by the assembly and disassembly of tubulins, components of the microtubule, and the change in ciliary membrane length. When the cell is non-dividing, the length of the primary cilium is consistent; however, when the cell enters mitosis, its length is shortened by the reabsorption of primary cilia into the cell body.121213141516 Recent studies have demonstrated that the presence and length of the primary cilium are associated with cell function.17181920 Furthermore, primary cilia control the cell cycle and cell migration, indicating that primary cilia play an important role in tissue injury and repair processes.22122 In the kidney, the primary cilium length is dynamically altered under the influence of not only physiological factors including renal flow and the cell cycle, but also pathological factors including reactive oxygen species (ROS), oxidative stress, and cytokines.78923242526 Recently we and others found that acute kidney injury (AKI) and chronic kidney disease (CKD) change primary cilium lengths in the kidney epithelial cells.567891127 Although it is unclear whether the change in primary cilia length is caused by these diseases because of the lack of evidence using animals with regulated cilia length, accumulated data indicate that the length of the primary cilium is associated with the progression of diseases. Furthermore, we recently found that the primary cilia of kidney cells exposed to oxidative stress and ischemia-reperfusion injury (I/R) are shed off into the culture medium and urine, respectively.79 These findings suggest that the length of primary cilia in kidney epithelial cells and the presence of primary cilia in the urine may reflect the status of the kidney cells and are useful indications of kidney diseases, including AKI and CKD. This article will deal with the association of primary cilia length alteration with several kidney diseases including AKI.
PRIMARY CILIA IN THE KIDNEY EPITHELIAL CELLS
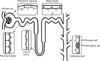
The primary cilium acts as a mechano- and chemo-sensor to detect renal fluid flow and composition, and mediates many extracellular signals into the cells in order to maintain renal function and nephron structure.318282930 In the kidney, a single primary cilium protrudes from the apical surface of renal epithelial cells above the microvillus.313233 The primary cilium, normally one cilium per cell, is found on the parietal cells of the Bowman's capsule and all kidney tubular epithelial cells, with an exception in the intercalated cells of the collecting ducts (Fig. 1).303233
Primary cilium lengths are diverse in the kidney cells, depending on the cell cycle and stage of kidney development; lengths may be short or non-existent in dividing cells, longest in redifferentiating cells, and medium-sized in mature cells.33034 In the adult mouse kidney, primary cilia length ranges between 2 and 7 µm in the tubular segments (the primary cilia lengths may not reflect precise sizes, because the data regarding primary cilia lengths were generated by kidney section staining), about 3 to 4 µm in the parietal cell, 2 to 4.5 µm in the proximal tubular cell, 2.2 to 3.7 µm in the distal tubular cells, and 1.6 to 2.6 µm in the collecting duct cell.56789112733 During the development of the kidney, the primary cilia are ~0.59 µm long and further lengthen to ~0.81 µm in S-shaped nephrons, finally extending to over 3 µm in human fetal and postnatal nephrons.34 Pazour et al. observed longer primary cilium length in the kidney tubular epithelial cells of 4 and 7-day-old pub mice when compared with those in mouse adult kidney; about 3.1±1.4 µm and 3.5±1.7 µm in the 4 and 7-day-old mouse kidney tubular epithelial cells, respectively, in scanning electron microscope (SEM) images.35 These studies indicate that non-fully differentiated cells have longer primary cilium when compared to fully differentiated cells, proposing that the cilia length reflects the cell status including the cell cycle. Furthermore, it indicates that the length of the primary cilia may be involved in the progression of kidney disease because, during the progression of disease, the kidney contains cells in diverse stages of cell-cycle.
ALTERATION OF PRIMARY CILIUM LENGTH
The primary ciliary length dynamically fluctuates during the cell cycle. The primary cilium disappears in the M phase and regrows in the G1, S, and G2 phases.132036 This growth of primary cilia is regulated by the assembly of axonemal microtubules in the core of primary cilia and membrane trafficking of ciliary membrane components from the intracellular producing site in mammalian cells.23537 In contrast, primary cilia shortening is regulated by the disassembly of microtubules and the release of ciliary proteins. The assembly and disassembly of primary ciliary proteins is controlled by outward anterograde (to the tip) and inward retrograde (to the cell body) transport of ciliary proteins.237383940 The intraflagellar transport (IFT) protein is generally recognized to be involved in these transportations. IFT consists of two multi-subunit complexes: IFT-A and IFT-B which comprise 6 and 16 subunits, respectively. IFT-A mediates both the retrograde and anterograde transport of ciliary proteins to shorten and elongate primary cilia, respectively. IFT-B mediates the anterograde movement of ciliary proteins to achieve the outward growth of primary cilia.24041 Recent studies have demonstrated that this IFT machinery is influenced by various factors including cdc14b, cyclic AMP activation of protein kinase A, hypoxia-inducible factor (HIF)-1α, cytokines, and mitogen-activated protein kinase signals.9162526273042 In addition, the organization of the actin cytoskeleton and the stability of the microtubule are also recognized as regulators of primary cilia length alteration.7164344
The shortening of primary cilia is associated not only with reabsorption but also with deciliation (shedding or disruption of primary cilia),79 although the shortening of primary cilia has been generally recognized as an absorptive process during the cell cycle.1256114245 Recent accumulated evidence reveals that primary length changes, including elongation and shortening, are adaptive responses to environmental stimuli, including I/R injury.56892028424546 Recently we found that kidney cell damage induces the deciliation of primary cilia in the kidney epithelial cells and the subsequent excretion of the primary cilia fragments into the urine.7914 Thus, a length change of the primary cilium has various clinical implications.
IMPLICATION OF PRIMARY CILIA LENGTH IN KIDNEY DISEASES
Epithelial cells of the diseased kidney reveal diverse ranges of primary cilia length during disease progression (Fig. 2),5,69111626 suggesting that the length of primary cilia reflects the status of diseases and that cilia length changes are influenced by various disease-associated factors, including inflammatory cytokines, ROS, HIFs, and regulators of the cellular cytoskeleton and cell cycle.78925262742 Ichii et al.24 recently reported that interleukin (IL)-1 receptor-like 2 (IL-1RL2), an IL-36α receptor protein, was expressed at the primary cilia of distal tubular epithelial cells in the contralateral kidney after unilateral ureteral obstruction (UUO), which induced the elongation of primary cilia.81124 They proposed that the elongation of primary cilia may increase the sensitivity of cells to urine flow and IL-36α.24 Verghese et al.25 reported that TNF-α does not affect cilia length in canine kidney tubular epithelial (MDCK) cells. These studies show that primary cilium length and cilium function are, at least in part, involved in inflammatory responses.
Recently we found that H2O2, a ROS molecule, induces the change of cilia length in the MDCK cells depending on its concentration; a high concentration of H2O2 induces shortening, whereas a low concentration of H2O2 induces elongation.79 This low-concentration H2O2-induced elongation of primary cilia was prevented by treatment with the extracellular signal-regulated kinase (ERK) inhibitor, U0126, indicating that ROS elongates primary cilia through the activation of the ERK signal pathway.9 Unlike low-concentration H2O2, high-concentration H2O2 causes the shortening of primary cilia in the MDCK cells, inducing increased primary ciliary proteins in the culture medium.79 This data indicates that ROS-stress induces the shedding of primary cilia, suggesting that the shortening of primary cilia in the injured kidney may be due to the deciliation. Rodríguez-Ribera et al.23 also found that reactive carbonyl compounds such as malondialdehyde, a product of oxidative damage, induce the loss of primary cilia in human kidney proximal tubule cells without the transit of cell to proliferation, that is, without reabsorption of primary cilia into the cell to re-enter the cell cycle. The ROS level in the kidney after I/R is dynamically altered; its level early after I/R is very high, whereas, later after I/R, this level is lower (but higher than the normal level) compared to the early state.4748 In the kidney, dynamic changes of primary cilia lengths may be associated with ROS and oxidative stress; primary cilia lengths were long in the kidney cells in the later phase after I/R, whereas they were short in the kidney cells in the early phase after I/R.569 This indicates that ROS and oxidative stress are associated with the alteration of primary cilia length and deciliation. Supporting this, studies have shown that antioxidant treatment affects cilia length changes in kidney epithelial cells subjected to I/R, unilateral nephrectomy (UNx), or hepatic I/R, accompanied by changes of ROS level, oxidative stress, and morphological recovery.78927
Furthermore, Verghesee et al.25 found that hypoxia and activation of HIF-1α induced by CoCl2 induce the elongation of primary cilia in MDCK cells. In this study, they suggested that the elongation of primary cilia may confer kidney disease resistance. It has been reported that the protection afforded by CoCl2 against acute and chronic kidney disease49 could be associated with the elongation of primary cilia and activation of primary cilia-based signaling.25 This data indicates that serial length changes in the kidney cilia length after injury may involve serial changes of ROS in the kidney tissue.
Primary cilia length changes are proposed as a repair and adaptive process of kidney cells to compensate for the increased renal burden after injury by coordinating signaling pathways.8162042 Recently, we found that the change in the expression of cell cycle regulator protein p21, a known protein cyclin-dependent kinase inhibitor 1, is involved in the primary cilia elongation in the tubule cells in the hypertrophic kidney following UNx.8 Lin et al.50 reported that p21 is inhibited in epithelial cells lacking primary cilia and that inactivation of Kif3a, a subunit of kinesin-II that is an essential protein for cilia formation, leads to the inactivation of p21. Basten and Giles,51 Basten et al.52 reported that, when cells enter the G0 phase, cells initiate ciliogenesis, whereas, when cells enter the proliferative phase, they lose primary cilia. These studies indicate that cell cycle regulators can control primary ciliogenesis and that primary cilia also affect the cell cycle in the kidney epithelial cells.
In addition, it has proposed that renal primary cilium defect and loss is associated with the repair of the damaged kidney.23 Saito et al.53 reported that the loss of primary cilia contributes to a switch from reversible to irreversible kidney injury in the UUO model, a progressive kidney fibrosis model, through the transition of non-canonical to canonical Wnt signaling in the tubular epithelial cells. Wnt signaling is activated at the nephron formation and repair process, whereas, it is suppressed in the adult kidney.54 Oh and Katsanis55 reported in the mouse UUO model that primary cilia emerged as principal modulators of Wnt signaling. Furthermore, a shortening of primary cilia is proposed to be a process that prevents harmful signals. Indeed, it reported that the loss of cilia is required for the remodeling of the injured epithelium.2056
Microtubule assembly and disassembly are critical for the alteration of primary cilia length,719455758 and are regulated by translational modifications including acetylation, tyrosination, glutamylation, and glycation of tubulins which comprise the microtubule.12154459 Pugacheva et al.58 reported that the reabsorption of primary cilia can be controlled by the deacetylation of axonemal microtubules. We also found that the shortening of primary cilia in the epithelial cells of acutely injured kidney cells following transient hepatic I/R is associated with the activation of the disassembly pathway and cytoskeletal disruptive pathway.7 Furthermore, we found that primary cilia length changes are associated with α-tubulin acetyltransferase and histone deacetylase 6 (HDAC6) expression.743 This indicates that translational modification is associated with primary ciliogenesis.
In all, accumulated evidence indicates that the process of primary cilia length change is closely linked to extracellular environments and intracellular cell signals which are associated with the pathogenesis of AKI and CKD, at least, suggesting that the control of ciliogenesis is an effective therapeutic strategy for a wide variety of kidney diseases and that the length of primary cilia reflects the progression of diseases.
CAN URINE PRIMARY CILIA PROTEINS AND TISSUE PRIMARY CILIA LENGTHS BE INDICATORS OF KIDNEY DISEASES?
Following exposure to pathogenic factors, the kidney is composed of various statuses of cells: lethal, sub-lethal, differentiated, and/or proliferated cells, depending on the degree of stress, exposure times, and so on.567891160 A kidney exposed to I/R injury is composed of diverse cells which have diverse primary cilia lengths, depending on the degree of injury and progression of injury followed by recovery (Fig. 2).5692742 According to time, primary cilia are shortened at early injury stages compared to normal-status lengths, are lengthened at the recovery stage, and gradually return to near their original sizes over time.569 In accordance with cell status, severely injured, dedifferentiated, and proliferated cells have short or even no primary cilia, whereas, redifferentiated and differentiated cells have long primary cilia.5679 Furthermore, the lengths of primary cilia are diverse through the tubular segments.5679 The kidney tubule segments have different susceptibilities to injury.616263 At the injury phase after I/R, the lengths of the primary cilia were shortened in the proximal tubular epithelial cells in the outer medulla but not in the cortex. The primary cilia length in the cortex and outer medulla were unchanged, whereas, the lengths of the primary cilia in the collecting ducts in the cortex and outer medulla were shortened. At the recovery phase after I/R, primary cilia lengths were longer than normal in all tubules, regardless of tubular segments.569
In the transient hepatic I/R-induced AKI mouse model, we also observed a similar pattern in the primary cilia length change.7 Furthermore, it has been reported that nephrotoxins, including cisplatin, shortened primary cilia in the kidney cell.42 These reports indicate that primary cilia length could provide information for defining the status of kidney cells and the fate of injured kidneys by accounting for the cell population of certain length ranges of primary cilia. In the human allograft kidney, Verghese et al.5 found that primary cilia lengths are dynamically changed after allograft, correlating with renal function. When renal function, as assessed by serum creatinine and glomerular filtration rate (GFR), was abnormal, that is in the acute tubular necrotic phase, primary cilia lengths were longer than normal, and thereafter, when renal function returned to the near-normal range, primary cilia lengths returned to the normal range.5 These animal and human studies indicate that the length of the primary cilia in kidney cells may reflect the status of the kidneys and that the length of primary cilia could be an index of AKI progression.
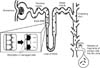
The primary cilia of kidney tubular epithelial cells project into the lumen of tubules. Upon cutting from the tubular and parietal cells, they drop into the ultrafiltrate, drain into the bladder, and are excreted in the urine (Fig. 3).79 Recently, we found increases of acetylated α-tubulin (ac-α-tubulin), the most broadly used marker of primary cilia, and ADP-ribosylation factor-like protein 2-like 1 (ARL13B), a protein located on the primary cilia to basal body, in the urine of mice at 4 hours and 24 hours, respectively, after 30 mins of kidney ischemia.79 However, we did not find GAPDH, an abundant intracellular protein, in the urine.9 This suggests that, although severe I/R injury in the kidney can lead to the detachment of tubular cells into the lumen, the deciliation of primary cilia is a rapid response by the cell to injury before the cells are completely separated from the tubules. The amounts of ac-α-tubulin in the urine were positively correlated with plasma creatinine levels.79 This indicates that the increase of primary cilia in the urine is associated with the injury of kidney cells and that the amount of primary cilia in the urine is correlated with injury levels. In addition, it indicates that the disruption of primary cilia may be started from the tip of primary cilia, since ARL13B is relatively more deeply and abundantly expressed near the cell body compared to ac-α-tubulin.
The shortening of primary cilia after kidney I/R injury is associated not only with the reabsorption of primary cilia for re-entry of cells into the cell cycle but also with the deciliation of epithelial cells.7942 In the partial hepatic I/R-induced AKI mouse model, we observed that primary cilia lengths were shortened in the kidney tubular epithelial cells 24 hours after hepatic I/R and that ac-α-tubulin levels in the urine were increased.7 The increase of ac-α-tubulin in the urine was observed 1 hour after hepatic ischemia when plasma creatinine level was not significantly increased.7 Furthermore, in the urine of patients subjected to hepatic resections, ac-α-tubulin levels markedly increased after surgery and then gradually decreased over time with hepatic functional recovery.7 The pattern of ac-α-tubulin expression in patient urine expression was similar to that of kidney injury molecule-1 (KIM-1) which is a well-known marker of kidney injury.764 Moreover, we found that the urine of kidney-transplanted patients contained ac-α-tubulin.7 This data indicates that the length of primary cilia in the kidney epithelial cells and the presence or increase of primary cilia proteins in the urine reflect kidney injury, suggesting that primary ciliary proteins in the urine could be very useful non-invasive biomarkers of kidney diseases.
CONCLUSION
Recent accumulated evidence strongly suggests that defective and abnormal primary ciliogenesis are associated with the pathogenesis of various kidney diseases and that the progression of these diseases affects primary cilia length changes, including deciliation due to the disruption. Moreover, human and animal studies have demonstrated that the detection of primary cilia fragments or ciliary proteins in the urine and the alteration of primary cilia length in the kidney cells could be useful for the diagnosis of kidney diseases; at least, AKI.
Although the above evidence implies that the control of primary ciliogenesis and deciliation may be useful for outlining treatment strategies for many kidney diseases, its use in the clinical setting, the exact molecular mechanisms involved, and sufficient evidence are still required. We believe that more broad and comprehensive studies of primary ciliogenesis and deciliation and the expression of cilia proteins in the urine will lead to improved approaches for diagnosing and treating kidney diseases.

 XML Download
XML Download