

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Global statistics estimate that approximately 25% of lung cancers are not directly attributable to smoking although cigarette smoking has been known to be the major cause of lung cancer (1). Moreover, only 10% of smokers develop lung cancer (2). These observations suggest that genetic variations related to the individual metabolism may determine susceptibility to lung cancer.
NAD(P)H:quinone oxidoreductase 1 (NQO1, previously called DT-diaphorase), a protein related to carcinogen metabolism, functions as a cytosolic flavoenzyme that catalyzes the electron reduction of substrate, to reduce oxidative stress (3). NQO1 catalyzes highly toxic quinones derived from tobacco smoking to less toxic hydroquinone analogues as a two-electron reductase (3, 4, 5). Moreover, cellular oxidation-reduction events controlled by this enzyme regulate the level of p53, a tumor suppressor (6). Because of their major role in cytoprotection and the inhibition of reactive oxygen generation in human tissue (7, 8, 9), the expression or mutation of the NQO1 gene has been considered to be potentially associated with the risk of cancer in the lung or other sites (10, 11, 12, 13, 14, 15, 16).
One point mutation in exon 6 of the NQO1 gene involves a C-to-T base pair substitution at position 609 of NQO1 cDNA, which codes for a proline-to-serine change at position 187 in the amino acid sequence of NQO1 protein. This NQO1 pro187ser polymorphism has been reported to be associated with greatly diminished levels of protein due to its accelerated degradation via the ubiquitin proteasomal pathway (17, 18). Moreover, in genotype-phenotype studies, cell lines and tissues obtained from persons with the homozygous mutant NQO1 genotype were reported to show deficient NQO1 activity (19, 20, 21, 22). Although several studies have reported a relation between the NQO1 pro187ser polymorphism and lung cancer risk, their results were found to be inconsistent probably due to small sample sizes, mixed populations, or disregard of polymorphisms in other sites of the NQO1 gene (10, 11, 12, 13, 14, 15, 23, 24, 25, 26, 27).
Therefore, in this study we estimated the effects of NQO1 gene polymorphisms on lung cancer risk using a haplotype-based approach in a relatively large sample.
MATERIALS AND METHODS
Study populations
A total of 616 Korean patients that were newly diagnosed with lung cancer at Chungbuk National University Hospital in Cheongju, Dankook University Hospital in Cheonan, and Inha University Hospital in Incheon between 2001 and 2003, and 616 controls without lung cancer that were individually matched to cases by age and sex were recruited. The exclusion and inclusion criteria applied were reported in a previous study (2, 28). Detailed information on diet, smoking habits, lifestyle, weight, height, medical treatment, and other socio-economic characteristics were collected by trained interviewers using a structured questionnaire. Venous blood samples of all subjects were collected into heparinized tubes and stored at -70℃ until used for DNA extraction.
Genotyping
In studies of single nucleotide polymorphism (SNP) based on limited numbers of subjects, low minor allele frequencies of SNPs may lead to null results, despite meaningful relationships between these SNPs and target diseases. Therefore, we first selected four NQO1 SNPs (rs689452, rs689453, rs1800566, and rs10517) which have more than 10% minor allele frequencies based on the SNP500Cancer database (http://snp500cancer.nci.nih.gov/home.cfm). Dan Stram's haplotype-tagging SNP program (tagSNPsv2.exe) was then used to determine the best set of haplotype-tagging SNPs. Based on this tagging SNP program, three SNPs (rs689452, rs689453 and rs1800566; min RSQ =0.9507) were selected from the above four and genotyped in 43 controls preferentially. One (rs689453) of these SNPs (rs689452, rs689453, and rs1800566) was excluded for further genotyping because it showed the same genotype (GG) in all of the 43 controls. Finally, two sites (rs689452, IVS1-27C>G; and rs1800566, Ex6+40C>T) were genotyped in all samples. Genotypings were performed completely blind of case-control status, and repeatability testing was conducted on 5% of the subjects, which resulted in a 99.6% concordance rate.
DNA was extracted from whole blood samples using a QIAamp DNA Blood Mini Kit (Qiagen, Valencia, CA, USA). SNP genotyping was performed using a single base primer extension assay (for rs689452) with the SNaPShot assay kit (ABI, Foster City, CA, USA) and using SNP-IT™ assays (for rs1800566) with the SNPstream 25K® System (Orchid Biosciences, Princeton, NJ, USA), as previously described (29). The designed primers were as follows: rs689452-forward, 5'-TCCTTTACAGACTGCAACTCC-3', rs689452-reverse, 5'-TCTGAGTGAGCCAGTACGATC-3', rs689452-SNP, 5'-TTTGCTGGTTGGTAATGGGTTTTCC-3'; rs1800566-forward, 5'-TGTGCTTTCTGTATCCTCAGAGT-3', rs1800566-reverse, 5'-ATTTGAATTCGGGCGTCT-3', rs1800566-SNP, 5'-TGCCCAATGCTATATGTCAGTTGAG-3'.
Statistical analyses
To identify whether each SNP site was on the Hardy-Weinberg equilibrium (HWE), the distributions of observed and expected genotype frequencies were compared using the chi-square test. When genotype data was not available for at least one of the two SNPs, the subjects with missing genotype were excluded in haplotype estimation. Individual haplotypes composed of two polymorphisms, IVS1-27C>G and Ex6+40C>T, were estimated from genotype data using the PHASE program (ver. 2.0.2). Linkage disequilibrium between two polymorphic sites was estimated as the relative disequilibrium (D') (30). Unconditional logistic regression was used to estimate odds ratios (OR) and 95% confidence intervals (CI) for the NQO1 polymorphisms in the lung cancer patients and control subjects. A probability level of 0.05 was used as the criterion for statistical significance, and SAS version 9.3 (SAS Institute Inc., Cary, NC, USA) was used for the statistical analysis.
RESULTS
The characteristics of the lung cancer cases and control subjects are shown in Table 1. Body mass index (BMI) and smoking were found to affect the OR of lung cancer (BMI, OR, 1.1; 95% CI, 1.06-1.14; and smoking, OR, 2.4; 95% CI, 1.86-3.09) while age, sex, and family history of cancer were not significantly different between cases and controls.
In the evaluation of the effect of the NQO1 genotypes (Table 2), we did not find any significant association with lung cancer risk after adjusting for BMI and smoking status, although the CC genotype of Ex6+40C>T showed a preventive effect for lung cancer risk as shown by its crude OR (crude OR, 0.7; 95% CI, 0.50-0.97).
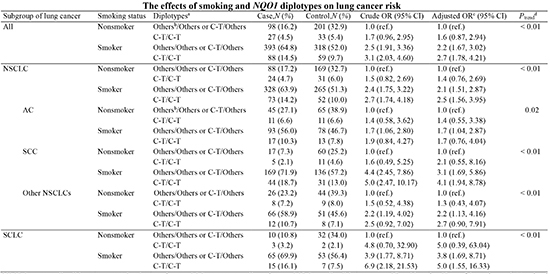
When we tested HWE of each NQO1 site, both cases and controls were found to be on the HWE (both IVS1-27C>G and Ex6+40C>T, P>0.05). In addition, the two sites were found to be strongly linked (D'=1, P<0.01). Therefore, we evaluated the effect of the NQO1 diplotypes for lung cancer risk and did not find any significant association with lung cancer risk after adjusting for BMI and smoking status (Table 3). Because smoking is a major risk factor of lung cancer and the NQO1 is related to the metabolism of tobacco carcinogens, we evaluated the effects of smoking in combination with the various NQO1 diplotypes. When smoking status and diplotypes were analyzed as categorical variables (Table 4), both smokers with the C-T/C-T diplotype and smokers without the C-T/C-T diplotype showed significantly higher risks of lung cancer than nonsmokers without the C-T/C-T diplotype (adjusted OR, 2.7; 95% CI, 1.78-4.21 and adjusted OR, 2.2; 95% CI, 1.67-3.02, respectively). In addition, the trend test showed an additive interaction of smoking with the NQO1 diplotype (Ptrend<0.01). Furthermore, when we estimated the combined effect of smoking and the NQO1 diplotypes on non-small cell lung cancer (NSCLC) and small cell lung cancer (SCLC) separately (Table 4), the additive effect of smoking and the NQO1 diplotype was found in both NSCLC and SCLC although the OR of lung cancer was greater in SCLC than in NSCLC. In the analyses of each NSCLC cell type (Table 5), the additive effect of smoking and NQO1 diplotype was found to be more apparent in squamous cell carcinoma (SCC) than in adenocarcinoma (AC) or in other NSCLCs, although all types showed similar trends in terms of the combined effect of smoking and NQO1 diplotypes on lung cancer risk.
DISCUSSION
NQO1 has been reported to be an important factor for carcinogen metabolism and cancer patient survival (3). However, previous studies have produced inconsistent results on the associations between lung cancer risk and NQO1 gene polymorphisms (10, 11, 12, 13, 14, 15, 26, 27). Because these inconsistencies may have been due to small sample sizes or disregard for other polymorphisms in the NQO1 gene, we used a haplotype-based approach in a relatively large number of subjects to increase statistical sensitivity. The two sites investigated in this study (IVS1-27C>G and Ex6+40C>T) were found to be linked strongly to each other. In connection with the strong linkage in our study, HapMap showed that these two loci exist as a haplotype block in the Asian population [http://www.hapmap.org/cgi-perl/gbrowse/gbrowse/], showing the reliability of our data and the importance of considering both of these loci.
In the present study, we evaluated the combined effect of smoking and NQO1 diplotype on lung cancer risk because smoking is the major factor in the development of lung cancer. Our study results showed that diplotypes of the NQO1 gene as well as smoking play an important role in the development of lung cancer and there is an additive interaction between smoking and NQO1 diplotype. These study results support the hypothesis that NQO1 has a shorter protein half-life in individuals with the T allele at Ex6+40C>T compared to individuals with the wild type C allele, and thus confers a higher risk of lung cancer development (17, 18, 19, 20, 21, 22). Moreover, in the present study, we used a web-based tool (Improbizer, http://www.cse.ucsc.edu/%7Ekent/improbizer/improbizer.html#NumMotifs) to identify whether or not the IVS1-27G>C polymorphism is located in a protein binding motif that has a potential to modulate NQO1 functionally although it is located in an intron. In the analysis using the web-based tool, this site was found to be located in the protein binding motif of NQO1 intron1, indicating the potential for the functional regulation of NQO1 protein activity.
In the present study, we also conducted subgroup analyses according to lung cancer cell type, because subtypes of lung cancer differ in many respects in terms of cellular origin, genetic and molecular changes, and clinical features. Trend analyses showed that the combined effect of smoking and NQO1 diplotype on lung cancer risk was statistically significant for all lung cancer cell types. However, our data also showed that this trend was more apparent in SCC. This more apparent relation with SCC is biologically plausible, because NQO1 protein may function as an oxidative stress inhibitor and SCC is more likely to be affected by tobacco smoke containing oxidative stress inducers than the other cell types of lung cancer (3, 31, 32, 33, 34, 35). Although SC-LC showed the largest OR of lung cancer in terms of NQO1 diplotype effect, the small number of nonsmokers with the C-T/C-T diplotype in SCLC patients and controls caused the confidence interval to broaden. Therefore, further studies are needed on a larger number of subjects to estimate the effects of the NQO1 diplotype on lung cancer risk in various types of lung cancer.
One of the weaknesses of the present study is that the control population was a heterogeneous mixture of healthy people and patients with a variety of different conditions that could be associated with NQO1 polymorphisms. This may possibly weaken our results even though we did not find any significant relationship between NQO1 polymorphisms and the development of diseases of the controls by our literature search (36, 37).
The present study suggests that diplotypes of the NQO1 gene play an important role in the development of lung cancer, and there is an additive interaction between smoking and polymorphisms in the NQO1 gene for the risk of developing lung cancer. However, we need to clearly understand the function of the NQO1 C-T haplotype in the development of lung cancer through further laboratory research.




 XML Download
XML Download