

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
The health benefits of regular physical activity for prevention of chronic diseases are well recognized. Exercise affects multiple physiological systems that over time leads to changes in body composition, skeletal muscle function, and exercise capacity [123]. Many previous experiments in animals have attempted to determine the biological and molecular mechanisms underlying the beneficial effect of physical activity on health [4]. However, many studies have used an involuntary animal exercise model, in which physical activity is induced by external stimuli. Involuntary exercise in animal models are not representative of normal exercise or physical activity in humans which is usually voluntary or motivated by positive stimuli. Thus, it is important to develop an alternative animal exercise model that avoids forced or involuntary physical activity and therefore can be translated into humans.
Regular exercise training protects against obesity as well as cardiac and skeletal muscle dysfunction [56]. Maximum endurance exercise capacity can be improved through re gular physical activity, which is determined according to environmental factors [789]. Evidence from animal exercise studies using different exercise modalities including treadmill running, swimming, ladder climbing, and standing indicate a strong association between physiological changes in body weight, cardiac function, and endurance exercise capacity [101112]. In forced exercise mo dels, exercise intensity can be maintained or increased using external stimuli, such as electrical shock and surgical ablation [11]. However, involuntary animal exercise models have several disadvantages including suppression of natural behaviours in the animal [13]. Therefore, it is important to develop and validate a voluntary animal exercise model, which results in physiological and functional changes such as enhanced endurance exercise capacity that is more representative to exercise and physical activity in humans. The development and validation of a voluntary animal exercise model is important to enable different exercise, diet or pharmaceutical interventions to improve exercise training-induced health benefits to be assessed in the future.
One recent study reported that involuntary stand-up physical exercises induced by electronic stimuli could cause skeletal muscle hypertrophy by enhancing protein synthesis in rats [12]. In this study, we determined whether voluntary stand-up physical activity (SPA) without electronic stimuli can effectively lead to similar benefits in terms of endurance exercise capacity and can help identify how primary exercise can be useful in involuntary exercise models. Thus, we assessed the effects of voluntary SPA on body composition, skeletal muscle tissue histology, running time, distance, echocardiography results, and hormone levels including irisin and corticosterone in rats.
Go to : 
METHODS
Animals and experimental design
Eight-week-old male Sprague-Dawley (n=12) rats (Orient Bio company, Daejeon, Korea) were maintained in an environmentally controlled facility (12:12-h light/dark cycle, 22℃). The rats were randomly assigned to two groups: sedentary control (CON, n=6) or voluntary SPA (n=6) for 12 weeks. All experimental procedures were approved by Inje Medical University Animal Care and Use committee for standard ethics in animal studies.
Voluntary stand-up physical activity
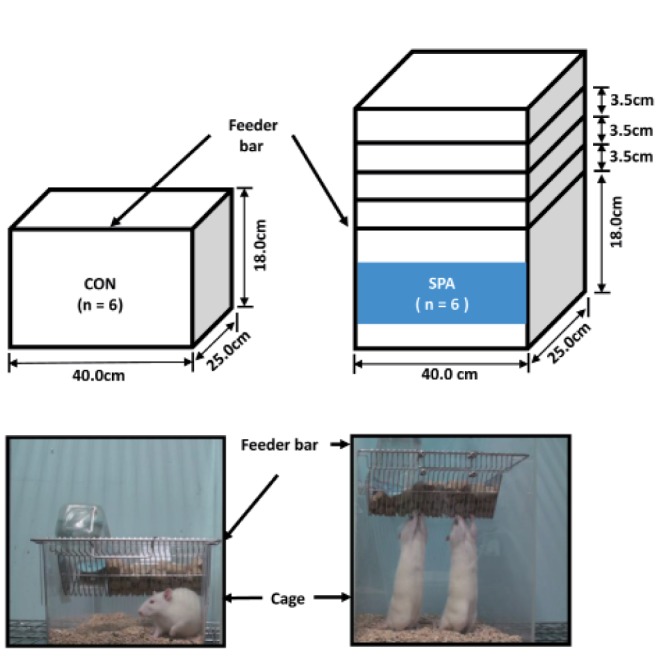
The rats were adapted to physical activity conditions (5 times/day, 60 min/times, with food maintained at a height of 1 stage: 21.5 cm) for 1 week (Fig. 1), without any stimulating factors, including electronic shock, light, and sound. As a result, the rats learned to stand, while consuming food and water. After the 1 week adaptation period voluntary exercise overload was increased progressively in three stages, using only body weight load, by increasing the height of the food cage up to a height of 28.5 cm. The voluntary SPA model was designed to be representative of daily voluntary standing type exercise and to mimic resistance exercise in humans.
Micro computed tomography
To determine the size of the lower limbs, we evaluated the lower limbs of the rats at 12 weeks with a SkyScan 1076 Micro computed tomography scanner (SkyScan, Kontich, Belgium) high-resolution system in vivo micro-computed tomography, as previous described [15]. A single representative CT image was taken for each group.
Echocardiography
To determine the cardiac function using transthoracic echocardiography results, rats were anesthetized with 2% isoflourane. We evaluated the complete 2-dimensional M-mode echocardiogram using the VIVID 7 dimension system (General Electric-Vingmed Ultrasound, Horton, Norway) and obtained images with a high spatial and temporal resolution by using a 10S transducer (5.5~12 MHz) [16]. The following data were recorded: heart rate (beats/min), stroke volume (ml/min), cardiac output (ml/min), fractional shortening (%), ejectional fraction (%), and left ventricle mass (mm). Ejection fraction was calculated as (enddiastole volume – end-systole volume) / end-diastole volume × 100%. Fractional shortening was calculated as (left ventricular diameter at diastole – left ventricular diameter at systole)/left ventricular diameter at diastole × 100%.
Grip strength
Grip strength was assessed in each rat using a grip strength meter (MK-380M; Muromachi Kikai Co., Tokyo, Japan) 3 days prior to the sacrifice [17]. Values were recorded as the average of three measurements.
Exercise endurance capacity
To determine the maximal exercise endurance capacity, treadmill running at 10 m/min for 5 min (grade 15°) was performed with an electronic shock stimulus and the speed was increased gradually at 2 m/min every minute, until the animals reached exhaustion. We evaluated exercise endurance capacity in terms of running time and distance [18].
Blood parameters
The concentrations of blood irisin (Phoenix Pharmaceuticals, Inc., Burlingame, USA) and corticosterone (Demeditec Co., Kiel-Wellsee, German) were measured using enzyme-linked immunosorbent assay kits.
Blood samples were taken from the heart and were drawn into heparinized tubes. The heparinized tubes were centrifuged for 10 min at 10,000 g to obtain plasma samples. All tissues were frozen in liquid nitrogen and blood samples were stored at –80℃ for next analysis.
Electron microscopy
To determine mitochondrial integrity, transmission electron microscopy was used. Samples from the gastrocnemius muscles were carefully dissected from the rats and analyzed as previously described [19]. Representative mitochondrial morphology of the gastrocnemius muscle was recored for each group.
Hematoxylin and eosin staining
The gastrocnemius muscles were embedded in liquid nitrogen and stained with hematoxylin and eosin, as previously described [20]. The number of nuclei of the gastrocnemius muscle was evaluated using Image J software (NIH, USA). A single representative cross sectional area of the gastrocnemius muscle was taken for each group.
Quantitative PCR analysis
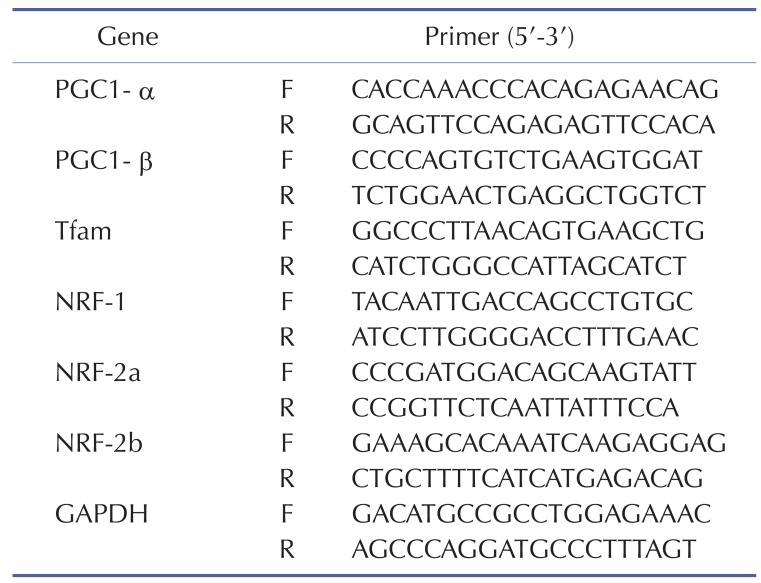
Total RNA from gastrocnemius muscle was extracted with Trizol (Invitrogen, Cergy Pontoise, France), and single-stranded cDNA was synthesized from 10 µg of total RNA with random hexamer primers (Applied Biosystems, Courtaboeuf, France). Real-time RT-PCRs were performed to measure PGC-1α, PGC-1β, Tfam, NRF-1, NRF-2a, and NRF-2b, as previously described [21]. Sequences of all primers are listed in Table 1.
Statistical analysis
All data are expressed as mean±standard error of mean (SEM). Changes in body weight within and between groups were evaluated using a two-way ANOVA [group (CON and SPA)×time (before and 12 weeks)]. If a significant change was noted in the group or time interaction, paired t-test was used for post hoc comparisons. Differences in other physiological and functional parameters between the CON and SPA group were evaluated by independent t-test (SPSS 22.0, Chicago, USA). Differences were considered statistically significant at p<0.05.
Go to :
RESULTS
Voluntary SPA reduced body weight but did not affect tissue weight or food intake
The reduction in body weight was significantly higher in the SPA group than in the CON group (Fig. 2A). Gastrocnemius and soleus weights were slightly higher in the SPA group than in the CON group, but the differences between the groups were not statistically significant (Fig. 2B, and 2C). The size of lower limb muscles were slightly bigger in the SPA group than in the CON group, but the differences between the groups were not statistically significant (Fig. 2D). Heart weight also did not significantly differ between the CON and SPA groups (Fig. 2E). To determine the changes in food intake, we measured food intake every week. There was no significant difference in food intake between groups (Fig. 2F).
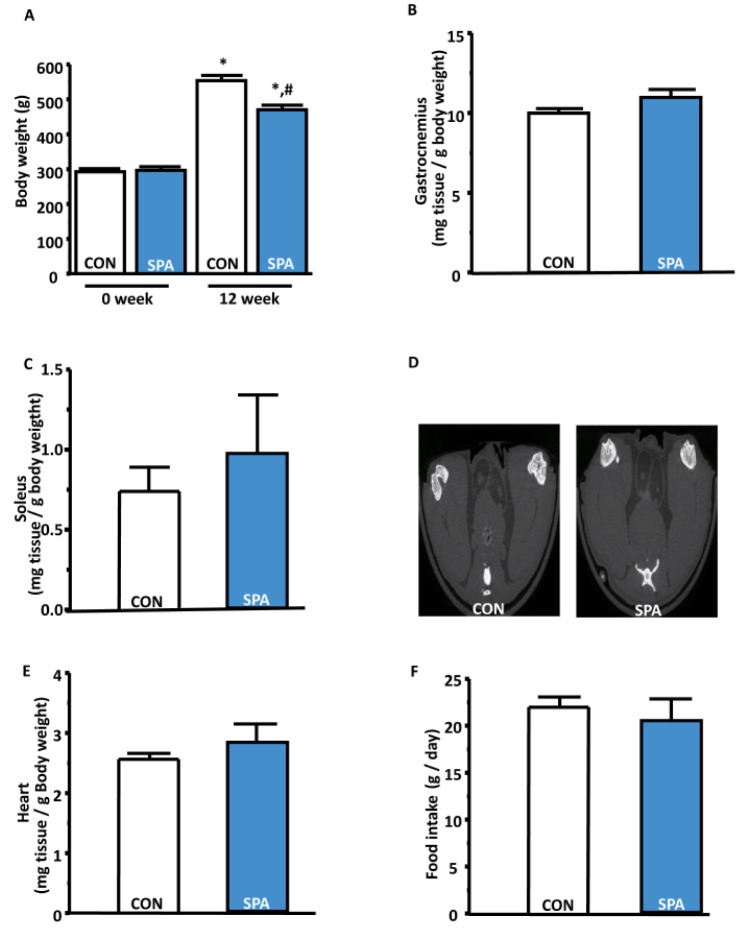 | Fig. 2The changes in body weight and organ tissues for a period of 12 weeks in the CON and SPA groups.(A) Body weight at 0 week and 12 weeks between the CON and SPA groups. (B) Gastrocnemius/body weight ratio in the groups. (C) Soleus/body weight ratio in the groups. (D) Lower limb muscle size that is only one representative CT image for each group. (E) Heart/body weight ratio in the groups. (F) Food intake/day ratio in the groups. The error bars indicate standard error of the mean (SEM) (*p<0.05; before vs after; #p<0.05; CON vs SPA). CON, control group; SPA, voluntary stand-up physical activity group.
|
Voluntary SPA did not change cardiac function, as determined via echocardiography
Increased physical activity typically is associated with improved cardiac function, including heart rate, left ventricle mass, cardiac output, ejection fraction, and fractional shortening [22]. However, no statistically significant differences in heart rate, stroke volume, cardiac output, fractional shortening, ejection fraction and LV mass were detectable between the CON and SPA groups in the present study (Fig. 3A-F).
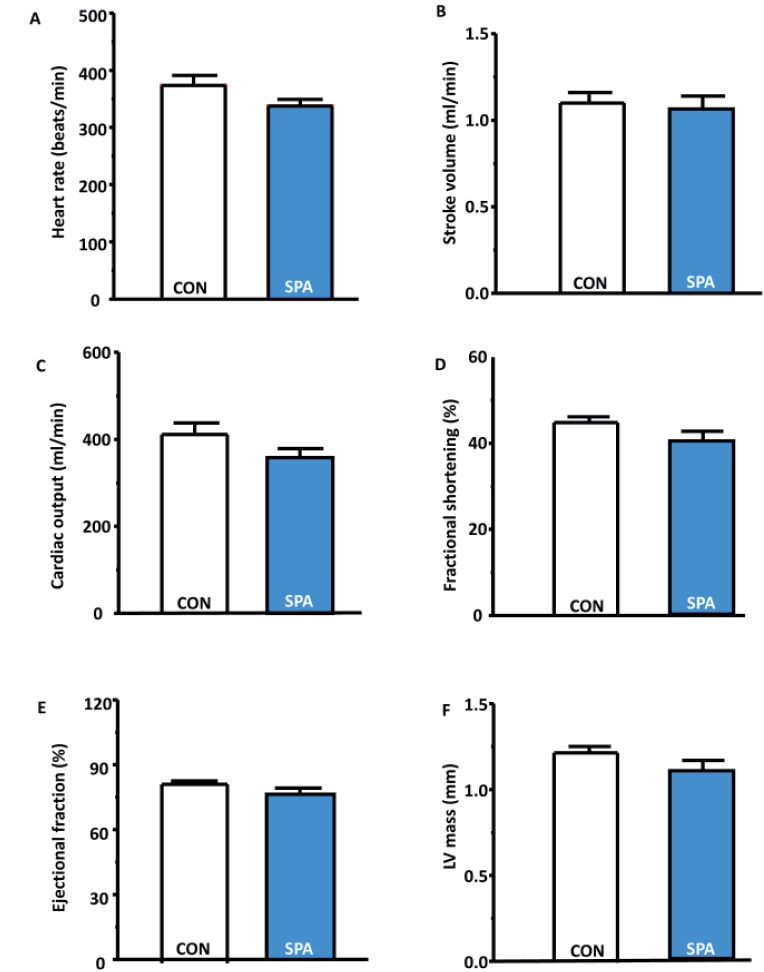 | Fig. 3Cardiac function as measured through echocardiography at 12 weeks in the CON and SPA groups.(A) Heart rate (beats/min) in the groups. (B) Stroke volume (mL/min) in the groups. (C) Cardiac output (mL/min) in the groups. (D) Fractional shortening (%) in the groups. (E) Ejection fraction (%) in the groups. (F) LV mass (mm) in the groups. CON, control group; SPA, voluntary stand-up physical activity group; Lv, left ventricle.
|
Voluntary SPA improved grip strength and endurance exercise capacity
The grip strength, as measured using a grip strength meter, increased significantly in the SPA group compared to the CON group (Fig. 4A). To determine changes in the exercise endurance capacity, we examined the difference in the acute exhaustive exercise test performance between the groups and observed that running time and distance were significantly higher in the SPA group than in the CON group (Fig. 4B, and 4C).
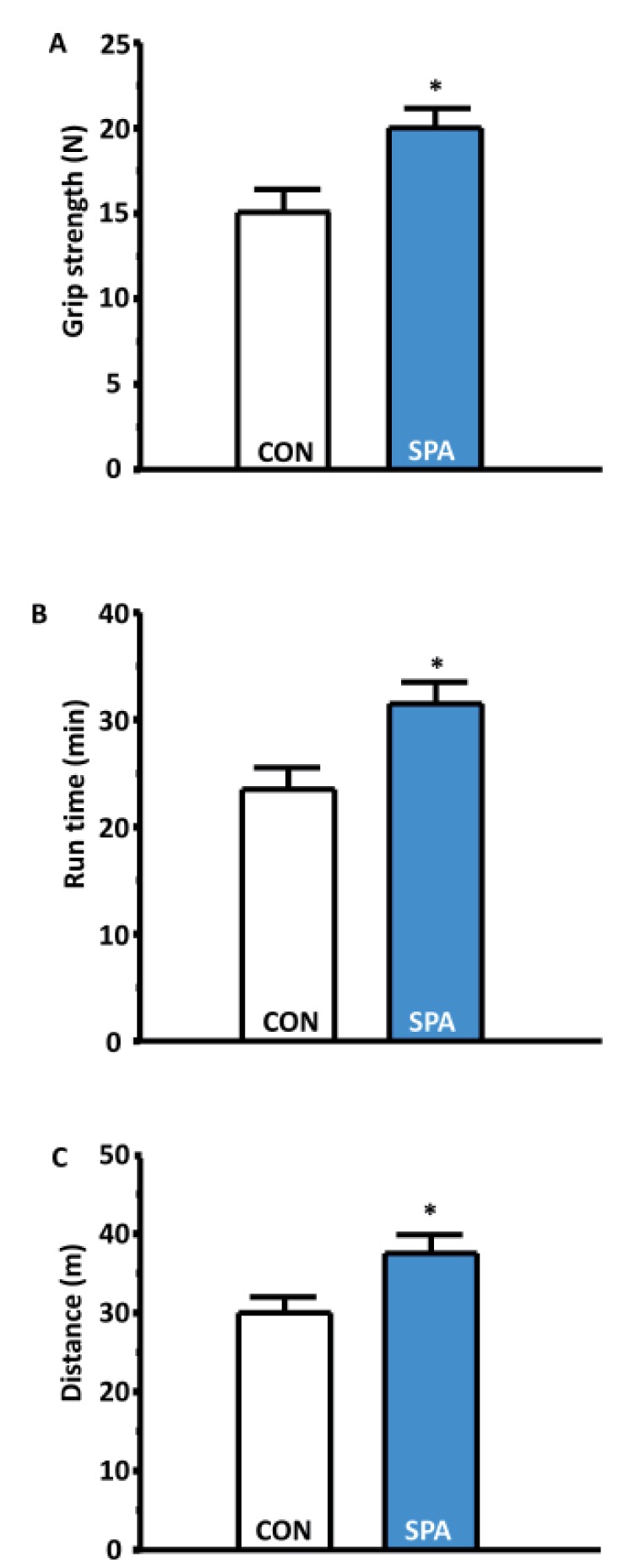 | Fig. 4Voluntary SPA shows enhancement of muscle strength grip strength and exercise capacity.(A) Grip strength (N) in the groups. (B) Run time (minutes) in the groups. (C) Distance (meters) in the groups. Error bars indicate standard error of the mean (SEM) (*p<0.05; CON VS SPA). CON, control group; SPA, voluntary stand-up physical activity group.
|
Effects of voluntary SPA on irisin and corticosterone levels
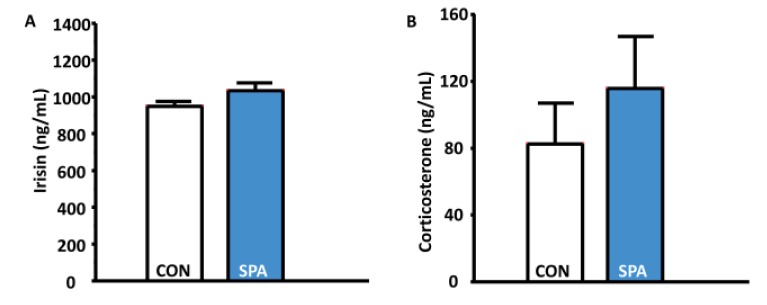
To support the changes in body weight and exercise endurance capacity, we determined irisin levels in the blood. In addition, we determined whether the corticosterone levels in the rats changed in the study. However, the levels of irisin and corticosterone, which tipically indicate stressful condition in animals were not significantly different between the CON and SPA groups (Fig. 5A and 5B).
Effects of voluntary SPA on mitochondrial abundance, mitochondrial biogenesis, nucleus, and muscle fiber phenotype in gastrocnemius in skeletal muscle
The SPA group showed slightly increased mitochondrial abundance, mitochondrial biogenesis, nucleus numbers, and muscle fiber size in the gastrocnemius than the CON group (Fig. 6A-E), but a statistically significant difference between groups was not seen.
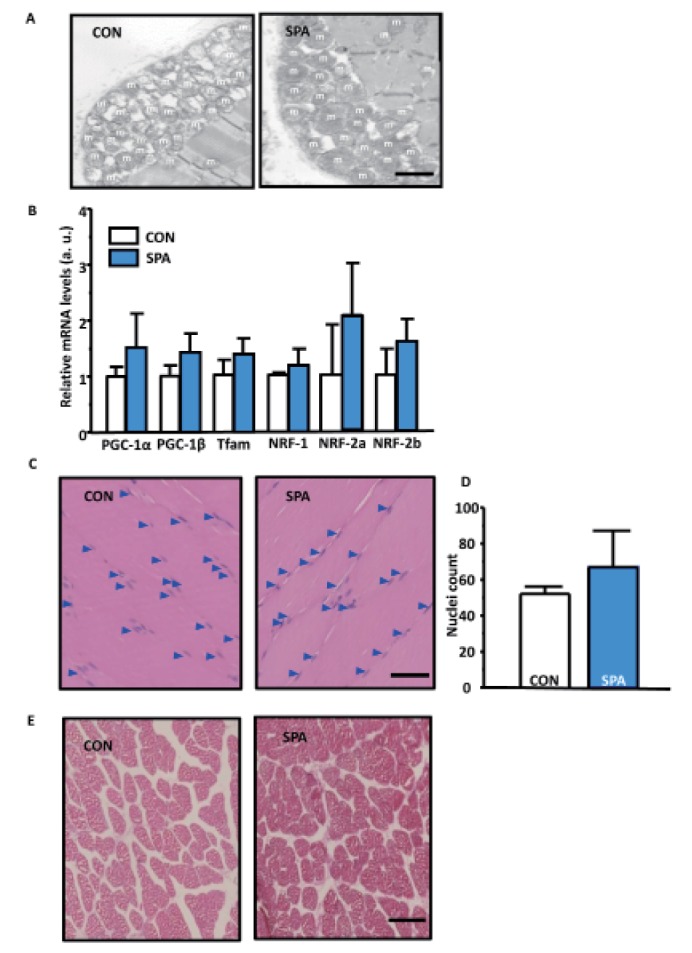 | Fig. 6Change in the histological morphology and mitochondrial biogenesis.(A) Mitochondrial abundant in the gastrocnemius in both groups. The arrow indicates the mitochondria. Scale bar indicates 500 nm. (B) Quantitative RT-PCR normalized to GAPDH mRNA. (C) Gastrocnemius hematoxylin and eosin (H&E) staining by using horizontal section in the groups. Scale bar indicates 40µm. An arrow indicates the nucleus. (D) Counted nuclei in gastrocnemius muscles. (E) Gastrocnemius H&E staining using cross section in the groups. Scale bar indicates 25 µm. m, mitochondria; CON, control group; SPA, voluntary stand-up physical activity group.
|
Go to :
DISCUSSION
In the present study, we first revealed that voluntary SPA without any negative external stimuli significantly decreased body weight, which was accompanied by significantly increased endurance exercise capacity and muscle strength after 12 weeks. However, voluntary SPA was not sufficient to improve cardiac function, skeletal muscle mass as well as the levels of irisin and corticosterone.
Increased physical activity results in the control of body weight, after considering energy supply according to energy consumption and expenditure associated with hormones [2324]. On the basis of these findings, we hypothesized that voluntary SPA would cause a decrease in body weight with a normal diet without stress. In the present study, we found that voluntary SPA was effective in decreasing body weight during 12 weeks (Fig. 2A). These findings are in agreement with previous study [25], which reported body weight reduction with voluntary wheel running compared with non-voluntary wheel running for 12 weeks, suggesting that the reduction in body weight may remarkably inhibit food intake and promote an increase in stress hormone levels. To confirm these findings, we assessed the food intake, and investigated the changes in corticosterone hormone levels. As expected, we observed no changes in the food intake or corticosterone levels between the two groups. These results indicate that the reduction of body weight seems to be a consequence of a response to voluntary physical activity without any external stimuli during voluntary SPA.
Generally, irisin as myokine, plays a role in body weight and total energy expenditure by activating brown adipose tissue cells during or after exercise [26]. In addition, it has shown that increased serum irisin levels results in the reduction of body weight [2627]. For these reasons, the increase of the serum irisin levels during exercise is considered a therapeutic strategy to reduce body weight. However, we showed that serum irisin levels were slightly higher in the SPA group than in the CON group, but no significant change was observed between the CON and SPA groups. These findings are in contrast to the results of many previous studies that showed that irisin levels were elevated as a reflection of the beneficial effects of metabolic response to exercise [2829]. Unlike the previous reports, the irisin levels in this study did not change significantly in response to exercise [3031]. Our design included a voluntary SPA protocol that inhibited body weight, but no changes in serum irisin levels were found. The present study demonstrated that voluntary SPA did not cause a change in irisin levels. Therefore, further studies on the mechanisms of irisin are necessary to determine whether voluntary SPA may be due to change in the body weight.
There are various exercise-induced protocols such as involuntary forced treadmill running, swimming, resistance type exercise, or voluntary wheel running that have effectively caused exercise metabolism adaptation in experimental animals [11]. These studies usually reported an increase in gastrocnemius and soleus weights in rats after 12 weeks of exercise training [32]. In the present study, although voluntary SPA showed the trend of increased weight in gastrocnemius and soleus muscles, these data did not reach statistical significant. In addition, we observed that the SPA group exhibited an increase in forelimb grip strength. This result led us to question whether the voluntary SPA enhances skeletal muscle fiber and nuclei or not. We showed that the SPA group showed slight increases in mitochondrial abundance, mitochondrial biogenesis genes, nuclei and skeletal muscle fiber size, but there were no changes in the gastrocnemius between groups. In a similarly designed study, stand-up exercise with electronic stimuli resulted in the rapid proliferation of myogenic satellite cells in atrophied muscles [12]. Our result may reflect that animals require a strong metabolic stimulus for skeletal muscle hypertrophy. Therefore, we suggest that SPA is seems more likely to be useful to augment grip strength.
Improvement in endurance exercise capacity in response to exercise is a well-recognized in cardiac and skeletal muscle function and is important to prevent chronic diseases [33]. As earier study, we assessed changes in echocardiography data during chronic physical activity stimulated by voluntary SPA. Although we observed an increase in endurance exercise capacity such as running time and distance in the SPA group compared with the CON group, echocardiography data did not show significant changes between the SPA and CON groups. These results indicate that voluntary SPA did not contribute to the enhancement of cardiac function. Recently, higher exercise endurance ca pacity has been associated with increased mitochondrial biogenesis, suggesting that exercise training may contribute to mitochondrial biogenesis [34]. In addition, we measured mitochondrial abundance in the gastrocnemius by studying electron microscopy images. We hypothesized in the present study that abundant and biogenetic increases of mitochondria would occur in the gastrocnemius, but we were unable to show statistical significant changes. We did not explore mechanisms of mitochondrial abundance, including oxidative metabolism and respiration and production of reactive oxygen species in response to exercise training. Further studies are necessary to address and understand the inhibition of body weight during exercise, with regard to mitochondrial function.
There are some limitations in the present study. First, the group size was quite small, with only 6 rats in each group. Second, we showed that voluntary SPA decreased body weight and increases endurance exercise capacity. However, we did not measure the level of energy expenditure during voluntary SPA. Third, we did not determine mitochondrial function and oxidative metabolism in the isolated mitochondria.
In conclusion, this is the first study to show that voluntary SPA decreased body weight and increased endurance exercise performance without any forced stimuli in animals, suggesting that it may be an alternative endurance exercise model in rats.
Go to :
 XML Download
XML Download