

 PDF
PDF ePub
ePub Citation
Citation Print
Print


TO THE EDITOR: L-asparaginase is an enzyme whose therapeutic effect has been well documented in children with acute lymphoblastic leukemia (ALL). The drug induces a relative asparagine deficiency that leads to the death of human lymphoblasts [1]. However, the mechanism of L-asparaginase-associated hepatotoxicity is not clear. There are few reports on the treatment of L-asparaginase-associated hepatotoxicity [234]. An experimental study conducted by Roesmann et al. [2] reported that when L-carnitine is administered in combination with L-asparaginase for steatosis, carnitine reduces the toxicity associated with L-asparaginase. Alshiekh-Nasany and Douer [3] reported that polyethylene glycol (PEG)-asparaginase related high-grade liver toxicity in adult patients could be rapidly and permanently reversed using the amino acid derivate L-carnitine. Al-Nawalki et al. [4] reported that histopathological macrovesicular steatosis was detected in 3 adult patients who developed L-asparaginase toxicity, and that the hepatotoxicity resolved after administration of carnitine and a vitamin B complex.
L-asparaginase is enzymatically active against glutamine, but with a significantly lower affinity against glutamine than L-asparagine [5]. L-glutaminase activity results in a reduction in the plasma glutamine level [6]. N-acetylcysteine (NAC) increases glutathione and antioxidant pools in hepatic cells, and thus increases the resistance of hepatic cell membranes [7]. Here, we describe a pediatric patient who developed severe hepatotoxicity accompanied by hyperamylasemia, hyperlipidemia, and elevated transaminase and bilirubin levels following the administration of 5 doses of L-asparaginase, and the parameters that were resolved following NAC infusion.
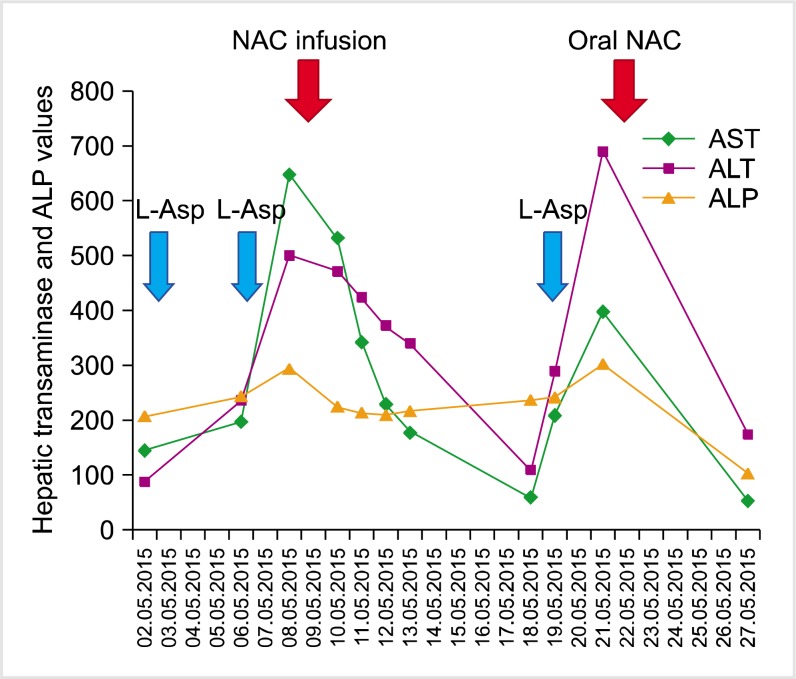
In the 2.5-year-old patient who was being followed-up after a diagnosis of pre-B ALL, induction therapy was initiated with a combination chemotherapy regimen comprising 60 mg/m2 prednisolone (oral) (days 1–33), 1.5 mg/m2 vincristine and 30 mg/m2 daunorubicin (days 7, 15, 22, and 29), and 5,000 µ/m2 L-asparaginase (E. Coli-Asp) (8 doses) (days 12, 15, 18, 21, 24, 27, 30, and 33). After the fifth L-asparaginase administration, the patient's laboratory results were as follows: alanine transaminase (ALT) 501 IU/L, aspartate transaminase (AST) 664 IU/L, gamma-glutamyl transferase (GGT) 1,297 IU/L, alkaline phosphatase (ALP) 246 IU/L, total/direct bilirubin 7/6.09 mg/dL, total cholesterol 272 mg/dL, triglyceride 571 mg/dL, and amylase 146 IU/L. The patient's clinical findings rapidly deteriorated, and abdominal distention and hepatomegaly were observed. Oral feeding was discontinued, and antibiotic (carbapenem and teicoplanin) and antifungal therapy (liposomal amphotericin B) was initiated. Tests for hepatitis A, B, and C, cytomegalovirus, Epstein Barr virus, parvovirus, and human immunodeficiency virus serology returned negative results. There was no growth in blood cultures. Abdominal computerized tomography revealed an increased liver size and density due to hepatosteatosis (Fig. 1). Drug-related toxic hepatitis was considered because of the progressively increased serum transaminase, bilirubin, amylase, and triglyceride levels. Liver biopsy could not be performed. The sixth dose of L-asparaginase was omitted. The NAC was started at a dose of 150 mg/kg for the first hour, 50 mg/kg for a subsequent 4 hours, and 100 mg/kg for the remaining 16 hours. Our patient received NAC infusion at a dose of 100 mg/kg/day over 4 days. The seventh administered dose of L-asparaginase was reduced by 50%. However, on day 2 of therapy, serum ALT was 689 IU/L, AST was 397 IU/L, and ALP was 302 IU/L. NAC therapy was re-started at an oral dose of 10 mg/kg daily and after the sixth day of NAC therapy, serum transaminase levels decreased to ALT 172 IU/L, AST 53 IU/L, and ALP 104 IU/L. After completion of induction chemotherapy, fasting cholesterol and triglyceride levels turned to normal. The administered therapies and changes in transaminase and bilirubin levels for induction chemotherapy are shown in Fig. 2. Hepatotoxicity was not observed during consolidation therapy, which consisted of high dose methotrexate. Consolidation followed a reinduction protocol, and Erwinia asparaginase was administered instead of E. Coli asparaginase. Although this protocol also used the same dose of vincristine and steroids as the induction protocol, hypertriglyceridemia was not observed. There was a temporary mild elevation of transaminases.
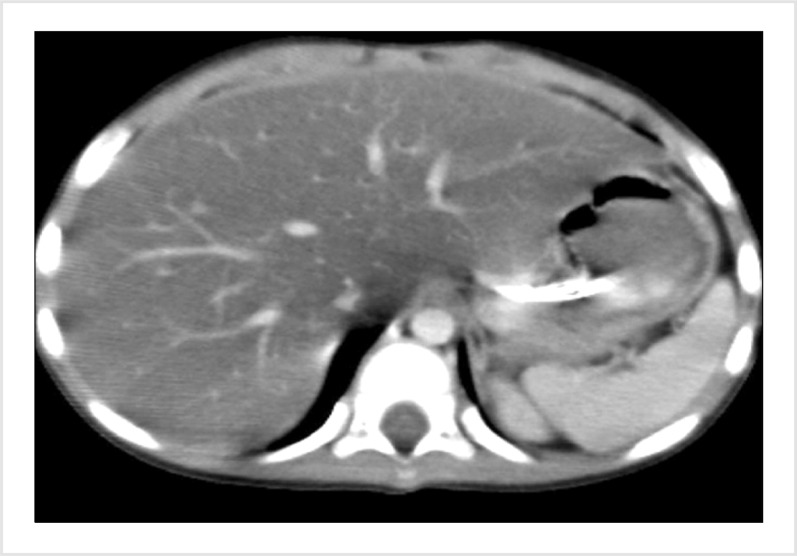
Liver dysfunction is a significant problem in patients receiving chemotherapy or immunosuppressive agents for hematologic malignancies. The most common cause of liver disease is drug toxicity. Furthermore, viruses can cause liver dysfunction. Hepatitis B is the most common of these viral agents [8]. Antibiotic therapy may lead to transient and mild transaminase elevation. Many drugs without antineoplastic effects such as allopurinol, ketoconazole, fluconazole, and ondansetron may cause hepatotoxicity but do not induce the fatty changes in the liver associated with steatosis [9]. Other agents administered as a part of the combination chemotherapy regimen, including daunorubicin and vincristine, are uncommon hepatic toxins that are not known to produce fatty changes. Fatty liver is a relatively common finding induced by L-asparaginase toxicity [10].
L-asparaginase induced hepatotoxicity appears to be related to oxidative stress, glutamine deficiency, and decreased hepatic protein synthesis, and subsequent impairment of beta oxidation in mitochondria [10]. As a result of the impaired mitochondrial beta oxidation, free fatty acids cannot be metabolized, and lactate and reactive oxygen radicals further increase and accumulate. The accumulated radicals lead to mitochondrial DNA damage, which results in disrupted aerobic metabolism in cells, and in turn lactic acidosis and triglyceride accumulation. The result is intracellular steatosis [11].
Glutathione is involved in the antioxidant defense mechanism against major reactive oxygen radicals. A small percentage (10–15%) of the total cell glutathione is present in the mitochondria. Despite this, mitochondrial glutathione plays a critical role in the maintenance of mitochondrial functions and in sustaining cell viability [12]. NAC is a glutathione precursor and acts by direct degradation of free oxygen radicals [7]. Previous studies have reported that NAC reduces lipid peroxidation and moderates the reduction in hepatic glutathione in non-alcoholic steatohepatitis patients [1314]. A study conducted by Baumgardner et al. [13] demonstrated that NAC is an effective hepatic antioxidant that eliminates the lipid peroxidation induced by non-alcoholic steatohepatitis, increases the reduction of hepatic glutathione, inhibits TNF-alpha production, and reduces inflammation, thereby significantly reducing hepatocyte damage.
Glutathione is an important intracellular antioxidant, which may be negatively affected by the reduction in glutamine levels that occurs as a result of L-asparaginase-associated inhibition of protein synthesis and L-glutaminase activity. In contrast, chemotherapeutics lead to increased oxidative stress and depletion of glutathione levels. The depletion and impaired synthesis of glutathione both result in a reduction in the glutathione pool. A study on mice demonstrated that in L-asparaginase-induced pancreatitis, glutathione levels were lower, and malondialdehyde, glucose, and triglyceride levels were higher in the group receiving asparaginase than in the control group and in the group receiving asparaginase concurrently with carnitine. Additionally, it was reported that in the group given asparaginase concurrently with carnitine, the glutathione, malondialdehyde, glucose, triglyceride, and amylase levels were closer to those of the control group than to those of the group receiving asparaginase [15].
Our patient's fasting triglyceride and cholesterol levels normalized, while the transaminase level showed a slight, temporary increase during reinduction chemotherapy with Erwinia asparaginase. These findings support that E. Coli asparaginase caused severe hepatotoxicity in induction chemotherapy. We recommend a NAC infusion dose of 150 mg/kg over 60 minutes, followed by 50 mg/kg over 4 hours, then 100 mg/kg over 16 hours in children with severe L-asparaginase hepatotoxicity. Maintenance treatment can be continued until clinical and laboratory findings are resolved. The decision to continue or cease NAC infusion should be at the clinician's discretion. This case demonstrates the utility of NAC supplementation for the treatment for severe hypertransaminesemia that developed 4 days after E. Coli asparaginase therapy.
 XML Download
XML Download