

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
Rearing animals specially in rodents have generally been treated by environmental enrichment (EE) to stimulate the effects of a complex combination of physical, cognitive, and social interaction [12]. EE is designed for enhancing social interactions, sensory and motor stimulation and learning and memory in a larger cage than standard cage, and this cage contains a variety of toys such as tunnels, platforms, and running wheels [34] (Fig. 1).
 | Fig. 1Representative finding of environmental enrichment.
An enriched environment was provided in a huge cage (86 × 76 × 31 cm3) containing various objects, such as tunnels, shelters, toys, and running wheels for voluntary exercise, and allowed for enriched social interaction (approximately 10–15 mice/cage) with a complex combination of motor, sensory, cognitive, and social stimulations.
|
Sensory, motor, and cognitive stimulation, interaction by EE are important elements in enhancing and adapting the neuronal system for the normal brain function [5]. Biochemical facilitation and morphological changes can promote functions in animals with brain injury [167]. Previous studies have been shown that the surrounding environment causes and stimulate many plastic responses in the adult brain, varying from biochemical parameters to dendritic arborization, gliogenesis, and neurogenesis [189]. EE significantly stimulates neuronal components such as cell body and size of nucleus, dendritic branching, dendritic spine density, and the amount of synapses [10111213].
Lately, researches have been focused on neural stem cells and neurogenesis because the neuronal replacement is the key point of brain reformation in the adult vertebrates [14]. Since EE supports the survival rate of the progeny cells of neuronal precursor in the hippocampus, the volume of the hippocampus expends in an animal model by increasing the number of neurons in the hippocampus [15]. An adult hippocampal neurogenesis is an active progression that can be induced by physiological and pathological causes. EE- or injury-induced hippocampal neurogenesis suggests common pathways of regulation by lesion and by environmental interventions [16].
EE can affect neural plasticity via overexpression of neurotrophic factors and neurotransmitter receptors [171819]. Several studies have shown that, in a brain lesion, brain-derived neurotrophic factor (BDNF), vascular endothelial growth factor (VEGF), insulin-like growth factor-1 (IGF-1), or fibroblast growth factor-2 (FGF-2) mediate the neurogenesis and angiogenesis promoting effects of EE [5710202122]. These growth factors are essential mediators of the effects of physical activities and EE in brain plasticity [23]. Moreover, EE have been reported as the treatment that can improve neurological functions in several animal models such as hypoxic–ischemia brain injury [7], and traumatic brain injury [24], Alzheimer’s disease [25], and Parkinson’s disease [2627]. Therefore, EE is considered as an advanced treatment to counteract cognitive and somatosensory deficits in neurological disorders with following brain injury [28]. Similarly, EE is known to have advanced effects on brain functions including the enhancement of motor and cognitive functions in normal and pathological states. The behavioral changes are related with these molecular changes [2930].
In this review, we will explain the brain plasticity and neurorestoration associated with molecular changes of neurotrophic growth factors such as BDNF, VEGF, IGF-1, FGF-2 and synaptic activity-regulating genes that occurs in interaction to EE.
BRAIN-DERIVED NEUROTROPHIC FACTOR (BDNF)
Function of BDNF is important in synaptic plasticity [3132] and previous studies report that its expression is changed following EE [533]. Physical exercise exposure has been shown to increase object recognition memory and to encourage BDNF expression in rats [3034], and the relations of EE and exercise on sensory gating and spatial memory has been formerly reported in rodents [3035]. In rodents, EE can improve memory and long-term potentiation (LTP) [4]. Another study described that BDNF is increased following EE, and this EE-induced BDNF goes with the enhancement of the object recognition memory and the improvement of LTP [5]. BDNF has been found to play a key role in the late phase of LTP, the best known cellular plasticity occurrence in the brain [363738], which is thought to be one of the plastic occurrence that underlies learning and memory [394041]. Especially, the hippocampus appears to be a specifically sensitive region to EE [42]: enriched animals show enhanced LTP [434445], improved learning and memory [464748], recovered behavioral functions after ischemia [49] and rescued memory deficits related to interruption of N-methyl-D-aspartate (NMDA) receptors [4546].
BDNF is also required for EE-mediated improvements of hippocampal neurogenesis [50]. Exposure of rodents to EE is an useful method for increasing both the adult hippocampal neurogenesis [155152] and the survival ratio of new-born neurons in the dentate gyrus [5354]. Since several studies described that neurogenesis and the survival of new-born neurons are connected to synaptic plasticity and memory function through BDNF/TrkB signaling [555657], it is likely that effects of EE on synaptic plasticity and cognition are enabled by these actions [5].
VASCULAR ENDOTHELIAL GROWTH FACTOR (VEGF)
The function of VEGF in the central nervous system (CNS) is comprehensive. Apart from its angiogenic effect in developmental and pathological angiogenesis [58], it has been described to have neuroprotective, neurotrophic and neurogenic effects [59606162]. Among these effects, angiogenesis is importantly associated with functional outcomes. At the same time, angiogenesis is essential for the repair of ischemic brain and functional recovery because it encourages blood flow and metabolism in the peri-ischemic region [2863]. Moreover, EE stimulated angiogenesis on cerebral cortex in an ischemic rat model. EE also encouraged the recovery of motor coordination and motor skills, and improved performances of learning and memory [28].
In addition, numerous studies have been reported that VEGF, as a direct mediator, improved cognition function in rodents [646566] and induced long-term synaptic enhancement in hippocampal neurons [67]. VEGF encourages hippocampus-dependent memory independently of its effects on neurogenesis and angiogenesis, by increasing the synaptic strength [6869]. Recent studies have been reported that short-term exposure to EE is also enough to alter synaptic plasticity and molecular markers related to the cognitive function [4]. EE-facilitated LTP was dependent upon NMDA receptors. These changes in synaptic plasticity occurred with raised levels of phosphorylated cAMP response element-binding protein (CREB) and VEGF [4].
Another study reported that hippocampal neurogenesis is responsive to VEGF expression by environmental inducements with enriched shelter and performance in a maze for the hippocampus-dependent test. This potential mechanism was confirmed by inhibiting VEGF expression by RNA interference, which entirely obstructed the environment induced neurogenesis [1664].
INSULIN-LIKE GROWTH FACTOR-1 (IGF-1)
IGF-1 has functions such as the control of cell proliferation, gliogenesis, neurogenesis, neuron survival, differentiation, synaptogenesis, and myelination in prenatal and postnatal CNS development [70717273]. Lately, neuroprotective effects of physical activities including exercise and EE support IGF-1 to regulate neuronal death [747576], the enhancement in hippocampal plasticity, and learning and memory in the adult [73777879]. The function of IGF-1 on both synaptic plasticity and hippocampal neuronal replacement seems mainly relevant to display such cognitive reserve. Like this, serum IGF-1 is necessary in the adult brain not only for both basal hippocampal neurogenesis and exercise and EE-induced increases of neurogenesis [22], but also for synaptic plasticity and cognition [23]. Recent study described serum IGF-1 has a direct role in cognition, although some long-term behavioral effects of exercise are IGF-1 independent at the same time [80].
Running also stimulates uptake of IGF-1 by certain groups of neurons enhancing electrical activity [76] and increases IGF-1 expression in the hippocampus [79]. Remarkably, EE has been shown to upregulate IGF-1 receptor gene in the adult hippocampus and sensorimotor cortex [81]. Especially, early exposure to EE could increase IGF-1 and BDNF levels in the brain, thus affecting hippocampal glucocorticoid receptor expression and stress response in adult brain [82].
In addition, IGF-1 increases astrocyte intercellular gap junctional communication [83], promotes dendritic arborization [84], regulates the rate of neurogenesis [23], and increases oligodendrogenesis [85]. Many studies described that learning and memory respond to IGF-1 after experience- or activity-induced events such as physical exercise and EE. All these actions, together with the above-mentioned evidences about the function of IGF-1 regulating diverse membrane channels, many neurotransmitter receptors, and neurotransmitter release, point to a control of critical aspects of neuronal excitability and, therefore, of the neuronal integrative capacities [2386].
FIBROBLAST GROWTH FACTOR-2 (FGF-2)
FGF-2 is existed throughout the CNS, and many previous studies reported that FGF-2 is important for the survival, reproduction, and differentiation of cells [878889]. The function of FGF-2 as a mediator on the brain is supported by demonstrations that FGF-2 is not only a solid proangiogenic factor [90] but also a neurotrophic factor which is generally produced by astrocytes in the adult brain [21919293]. Previous study reported that expression of FGF-2 was expressively raised in mice exposed to EE compared with exposed to standard cage, causing that the upregulation of FGF-2 might be a key factor in functional recovery after hypoxic-ischemic brain injury [7]. In addition, increasing FGF-2 protein by intraventricular infusions induced increased hippocampal neurogenesis [9495]. The studies showing a FGF-2 dependent neurogenic response in the dentate gyrus and subventricular zone (SVZ) from ischemia are according to a condition for injury and/or inflammation facilitating FGF2-mediated neurogenesis [72196].
The elevated expression of FGF-2 induced by EE improved neurogenesis and angiogenesis in animal models of chronic hypoxic-ischemic brain injury [721]. Physical activities including exercise and EE are interesting to speculate that these factors act at least in part by stimulating FGF signaling [97]. The previous study described that FGFs directly affected on hippocampal stem cells [94] and that the decreased neurogenesis with age is not only due to a decreased ability of stem cells to generate new neurons or respond to FGF signals, but also a decrease in the availability of ligands in their environment [97]. In addition, FGF-2 have a potent ability of chemotactic factor for endothelial cells [98], and it showed the function of regulating recovery from cerebral brain injury [99100]. FGF-2 improves sensorimotor deficits and to decrease infarct size following cerebral ischemic brain injury in adult rats [101], and neutralizing antibodies to FGF-2 blocks recovery from motor cortex lesions [102].
Since FGF-2 is initially essential for migration of neural progenitors [103], neuronal recruitment into the striatum may have been potentiated by increased FGF-2 [21]. Moreover, neurogenic repair processes mostly depend on FGF-2, which is largely expressed by reactive astrocytes in the injured brain [721]. Although both risky and beneficial activities have been attributed to reactive astrocytes, recent studies have shown functions for these cells in limiting inflammation and protecting neurons and oligodendrocytes in brain lesions [104105]. EE has been shown to increase overall expression of astrocyte marker, glial fibrillary acidic protein (GFAP) similar to the pattern of FGF-2 released from activated astrocytes [106], which suggests that astrocyte activation can mediate both neurogenesis and angiogenesis, consequently resulting in functional improvement through a FGF-2 dependent mechanism [721] (Fig. 2).
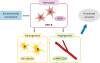 | Fig. 2Brain plasticity and neurorestoration by environmental enrichment.
Environmental enrichment has been shown to increase overall expression of astrocyte marker GFAP similar to the pattern of FGF-2 released from activated astrocytes, which suggests that astrocyte activation can mediate both neurogenesis and angiogenesis, consequently resulting in functional improvement through a FGF-2 dependent mechanism.
GFAP, glial fibrillary acidic protein; FGF-2, fibroblast growth factor-2.
|
SYNAPTIC ACTIVITY-REGULATING GENES
Experience-driven changes in brain include both modifications of the synaptic connectivity of the circuits in a local synapse-specific manner [107], and the induction of activity-dependent gene expression [108]. An increasing number of molecules and genes have been involved in activity-induced plasticity [23]. Previous studies [1109] reported that the expressions of genes related with neuronal structure, synaptic plasticity, and transmission were significantly changed by short-term and long-term exposure to EE. When microarray-based determination of gene expression patterns has been used to find pathways that are affected by EE [1109110], and recent study [1] specifically described that the gene sets expression related with various synaptic changes including chemical synaptic transmission, postsynaptic neurotransmitter receptor binding and G protein activation, kainate receptor activation, the opening of the presynaptic calcium channel, and acetylcholine release were up-regulated, while those associated with neurotransmitter transporters such as dopamine transporters were down-regulated [19].
Above all described things, G protein activation is important for transducing transmitter attachment into postsynaptic responses [111]. Specially, synaptic plasticity is controlled by G protein coupled receptors whose signal pathways aim the NMDA subtype of excitatory glutamate receptor in hippocampus [112]. The recent study [1] exhibited that the expression of G protein-coupled purinergic receptor P2RY12 increased in the hippocampus, where the proteins were mainly spread, regulating the signaling functions in neurotransmission and regeneration [113114]. It is probable that the upregulation of P2RY12 might be related with improved memory function [1].
Activation of kainate receptor, a non-NMDA glutamate receptor, also has a function in synaptic plasticity, affective the postsynaptic cell firing and the presynaptic neurotransmitter release underlying regulation of excitatory synaptic transmission in the hippocampus [115]. The molecular mechanisms underlying these alterations in synaptic features have been traced to NMDA receptor-dependent synaptic plasticity [116] and rises in postsynaptic [Ca2+] [117]. In this way, it is remarkable that the physical stimulation including EE is a physiological inducement delivering the brain with peripheral trophic sustenance [123].
In addition, EE-directed improvement of motor and cognitive functions is not only related to growth factors but also related to the modification of synaptic activity-regulating genes [1]. These genes are related with postsynaptic receptor activity, and the genes are modulated in the presynaptic neurotransmitter transporters that potentially leading to the more efficient use of neurotransmitters and enhanced synaptic plasticity. For examples, dopaminergic receptors are up-regulated in the postsynaptic area, and on the other hands, dopamine transporters are down-regulated in the presynaptic area, consequently resulting in the increase of the dopamine neurotransmitter in the synaptic level [119].
CONCLUSION
EE enhanced neural plasticity such as neurogenesis, gliogenesis, angiogenesis, and neurobehavioral functions. Additionally, EE can affect neural plasticity via overexpression of various neurotrophic growth factors such as BDNF, VEGF, IGF-1, FGF-2 and synaptic activity-regulating genes such as Na+/Cl−-dependent neurotransmitter transporters including dopamine transporter. Therefore, EE is considered as an advanced treatment to counteract neurological disorders with cognitive and somatosensory deficits following brain injury. We reviewed the brain plasticity and neurorestoration associated with molecular changes of neurotrophic growth factors such as BDNF, VEGF, IGF-1, FGF-2 and synaptic activity-regulating genes in interaction to EE. Taken together, these findings suggest that EE has beneficial effects on the brain plasticity and neurorestoration.
 XML Download
XML Download