

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Asthma is a chronic respiratory disease mediated by a wide range of environmental and genetic factors.1 It is characterized by coughing, wheezing, bronchoconstriction, airway hyperresponsiveness, or airway remodelling. Due to its multifactorial nature, several studies have attempted to elucidate the genetic background of asthma pathogenesis.23456
The Hippo pathway is highly conserved from Drosophila melanogaster to mammals and regulates organ size through promoting apoptosis and inhibiting cell proliferation in the embryonic stages of development.7 It is still not exactly known how the pathway is regulated by organ size; however, it has been proposed that FRMD6 (also known as Willin) influences the activity of the Hippo pathway by turning on the central kinase cascade.8 The members of this signaling cascade, MST1/2 and LATS1/2 with scaffold proteins SAV1 and MOB1, respectively, phosphorylate one another to inhibit YAP1/TAZ, the main effectors of the pathway.9 YAP1 and TAZ are transcriptional coactivators that bind to transcription factors, when active, such as TEAD, SMAD, or TP73, to regulate the expression of anti-apoptotic (e.g., BIRC5), or apoptotic genes.101112 Recently, it has been shown that YAP1 is also widely expressed in respiratory epithelial cells of the embryonic and mature lung and that Hippo/YAP1 signalling regulates epithelial cell proliferation and differentiation, and play an important role in embryonic lung maturation and postnatal airway homeostasis.13 Furthermore, in mice it has been demonstrated that YAP is dynamically regulated during regeneration of the airway epithelium following lung injury, suggesting a possible role of Hippo/YAP1 signaling in the pathogenesis of acute and chronic lung diseases.14
Previously, it has been found that genetic variations in FRMD6, an upstream activator of the Hippo pathway, were associated with asthma.15 Furthermore, genetic variations in the BIRC5 gene (also known as survivin), which is one of the target genes of YAP1 (Fig. 1), influenced the susceptibility to the disease.16 In addition, the sputum mRNA level of FRMD6 was significantly lower, while that of BIRC5 was higher in asthmatic patients than in healthy controls.1516
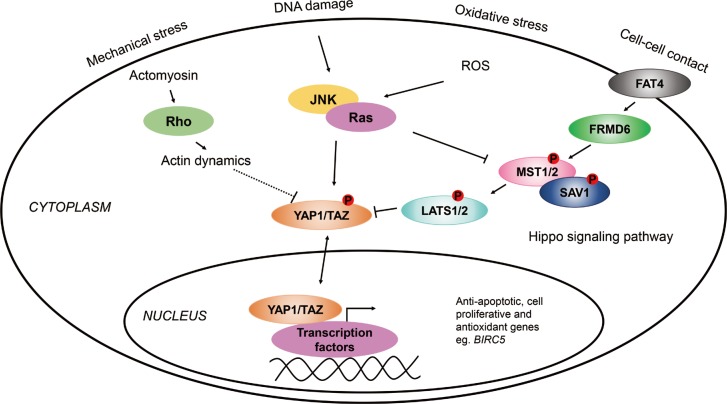
Based on these data, we hypothesized that the Hippo/YAP1 pathway may play a role in the pathogenesis of asthma. The main aim of our study was to investigate the members of the Hippo pathway and compare their gene expressions in the induced sputum of asthmatic patients and healthy controls. Moreover, we also determined whether genetic variations in the YAP1 gene can influence the susceptibility of asthma or subgroups of asthma.
To evaluate the role of the Hippo/YAP1 pathway and one of its target genes, BIRC5, as a system in asthma and allergy, we also analyzed our data with the BN-BMLA method, in order to provide more information on the relationship between genetic variations in the YAP1, FRMD6, and BIRC5 genes, asthma susceptibility, as well as other types of variables, e.g., allergy, eosinophils, immunoglobulin E (IgE), and asthma phenotypes.
MATERIALS AND METHODS
Subjects
Gene expression analysis was carried out using the induced sputum of 18 asthmatic patients and 10 healthy controls. All the study participants completed a detailed, pre-edited questionnaire. The recent Global Initiative for Asthma (GINA) guidelines (www.ginasthma.org) were used to diagnose asthma by a respiratory medicine specialist. The evaluation of asthma severity was done at the time of acquisition of induced sputum samples from the patients based on patient history, including number of exacerbations per year, lung function test results, medical treatment applied, and response to medication. The asthmatic patients were divided into 4 severity groups; however, Global Initiative for Asthma 1, 2 (mild) and GINA 3, 4 (moderate-severe) were aggregated because of the small number of patients. Of the 18 asthmatic patients, 14 regularly used inhaled corticosteroids (ICS): <500 µg/day beclomethasone dipropionate (BDP) or equivalent (n=5), 500-1,000 µg/day BDP or equivalent (n=7), and >1,000 µg/day BDP or equivalent (n=2); while 4 were considered steroid naive. Controls were healthy volunteers with no previous history of asthma or any airway conditions. All subjects participated in a lung function test (PDD-301/S; Piston Inc., Budapest, Hungary) and were assessed for fractional exhaled nitric oxide (FENO) levels (NIOX MINO; Aerocrine, Solna, Sweden). All the healthy patients had a normal lung function and had no respiratory tract infection 4 weeks prior to the analysis. A skin prick test was also performed for common allergens in order to test the presence of atopy, which is a genetic predisposition to develop allergic rhinitis, atopic dermatitis, and/or asthma. The participants' sex, age, smoking habits, and allergic statuses were compared between cases and controls, and between severity groups, but no statistical significance was found.16
Table 1 shows a summary of this study population.
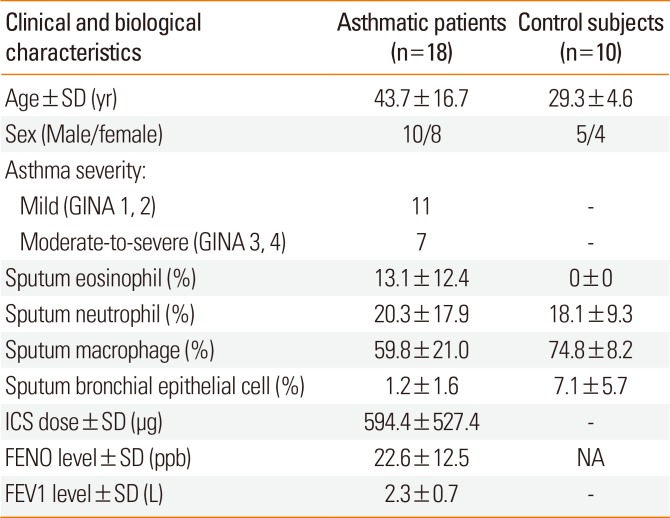
Table 1
Detailed characteristics of subjects participating in sputum analysis
![]()
The genotyping analysis included 1,233 unrelated individuals, out of which 522 were asthmatic children (mean age±standard deviation [SD], 10.20±5.31 years; 218 males and 194 females), and 711 healthy controls (mean age±SD, 14.00±11.22; 429 males and 282 females). The patients were all Hungarian (Caucasian), from which about 5%, based on state population data, were of Gypsy origin. Asthma was diagnosed based on the GINA guidelines as previously described. Atopy was identified by a positive skin prick test and/or positive total or specific serum IgE levels. The total and specific serum IgE levels were tested with the 3gAllergy blood tests in the Immulite 2000 Immunoassay system (Siemens Healthcare Diagnostics, Tarrytown, NY, USA). The resulting serum IgE levels were either normal or high based on age-specific ranges (kU/L).
Different types of asthma were defined in patients, except for 131 subjects (25%), where not enough information was available on asthma phenotypes; hence altogether, 391 asthmatic children were involved in the phenotype analysis. Asthma was divided into allergic and non-allergic asthma subtypes. If asthma was not provoked by allergens but allergy was also present, the allergy types were marked. In allergic patients, depending on the types and quantities of allergens, subgroups of indoor, outdoor, or inhalative allergic phenotypes have been created. If asthma exacerbation was provoked by exercise in the medical history of the patients, asthma was categorized as exercise-induced one. If the onset of asthma or asthma exacerbations have been associated with an infection-related acute respiratory illness, asthma was classified as viral-induced one. Non-atopic patients with viral-induced asthma phenotype composed the non-allergic asthma subgroup.17 Indoor allergens included dust mites, mold, pet dander, and cockroaches, whereas outdoor allergens consisted of different types of pollen. Eosinophil cell counts from blood were measured with the Coulter AXM analyzer, of which the normal relative range was between 1%-6%, and the normal range of absolute eosinophil count was between 50-200/µL. The patients had no change of therapy before the blood samples were taken. Neither did they have exacerbations or respiratory infections for at least 4 weeks prior to the blood test. The detailed characteristics of the asthmatic patients participating in the gene association study are presented in Table 2.
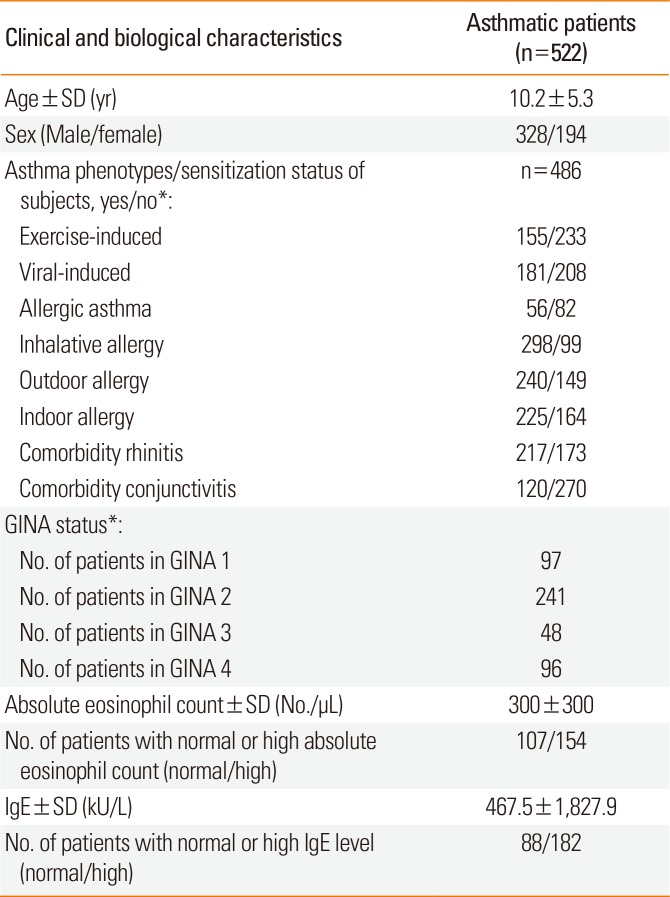
Table 2
Detailed characteristics of asthmatic patients participating in SNP analysis
SNP, single nucleotide polymorphism; SD, standard deviation; GINA, Global Initiative for Asthma; IgE, immunoglobulin E.
*Data are available on a limited data set only. Normal absolute eosinophil count is <200/µL and high absolute eosinophil count is ≥200/µL. Normal IgE level is <200 kU/L, high IgE level is ≥200 kU/L.
![]()
The control children were from the Orthopaedic Department in Budai Children's Hospital or from the Urological Department from Heim Pál Hospital, both in Budapest. The controls had no symptoms of asthma or any airway conditions, nor any need for medication.
Written informed consents were signed by all patients or by their parent/guardian. The study was done according to the Declaration of Helsinki and approved by the Ethics Committee of the Hungarian Medical Research Council.
Sputum induction
Induced sputum was used for gene expression assays and Western blot analysis. Sputum samples were taken and prepared as previously described by Ungvári et al.16
RNA isolation and cDNA production
RNA was isolated successfully from the induced sputum samples of 18 patients and 10 control subjects with the Qiagen Mini RNeasy Kit according to the manufacturer's instructions (Qiagen, Germantown, MD, USA). The cDNA used in the gene expression analysis was produced with the High Capacity cDNA Reverse Transcription Kit from Thermo Fisher Scientific (Thermo Fisher Scientific, Waltham, MA, USA).
Gene expression analysis
Real-time quantitative polymerase chain reaction (PCR) was performed on LATS1, LATS2, MST1, MST2, SAV1, YAP1, TAZ, and β-actin genes using the 7900HT Fast Real-Time PCR System (Thermo Fisher Scientific) in which β-actin was used as an endogenous control, and all results were normalized to it.
DNA isolation and genotyping
Genomic DNA was isolated from the whole blood samples of 1,233 individuals using the QIAamp blood DNA midi kit (Qiagen) or the iPrep PureLink gDNA Blood kit on the iPrep Purification Instrument (Invitrogen, Carslbad, CA, USA). KBiosciences Competitive Allele-Specific PCR (KASP) version 4.0 genotyping assays were used (LGC Genomics, Berlin, Germany) to genotype 14 single nucleotide polymorphisms (SNPs) on the YAP1 gene (Table 3) according to the manufacturer's instructions. PCR reactions were carried out using the 7900HT Fast Real-Time PCR System (Thermo Fisher Scientific).
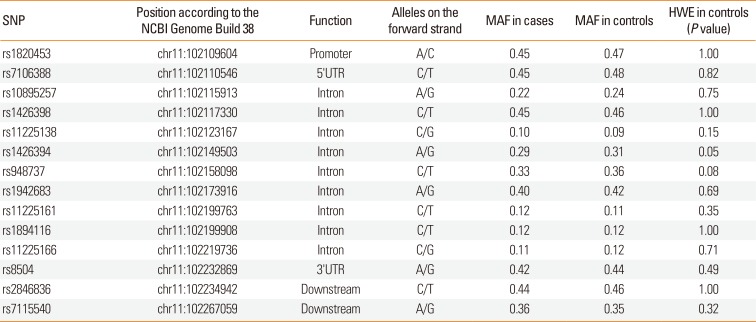
Table 3
Description of selected SNPs and results of the genotyping
![]()
Western blot analysis
Western blot analysis was carried out with human induced sputum samples. After sample preparation, cells were lysed in RIPA lysis and extraction buffer (Thermo Fisher Scientific) supplied with Halt protease inhibitor (Thermo Fisher Scientific) at 1x final concentration and centrifuged at 14,000×g at 4℃ for 15 minutes. Total protein content was determined by the Pierce BCA protein assay kit (Thermo Fisher Scientific) according to the manufacturer's instructions. Samples were loaded on 4%-20% Tris-glycine precast gels (Thermo Fisher Scientific) and then blotted onto the Immun-Blot PVDF membrane (Bio-Rad Laboratories, Hercules, CA, USA). After blocking, the following antibodies were used. The primary antibodies were anti-FRMD6, anti-YAP1, and anti-GAPDH (Abcam, Cambridge, UK). The secondary antibodies were polyclonal donkey anti-rabbit IgG HRP (Abcam) and polyclonal goat anti-mouse immunoglobulins HRP (Dako, Glostrup, Denmark). The membrane was treated with Pierce ECL plus substrate (Thermo Fisher Scientific) according to the manufacturer's instructions, and bands were visualized on a standard X-ray film (Kodak, Rochester, NY, USA).
Frequentist statistical analysis
For sputum analysis, normalized gene-expression levels were compared by the Mann-Whitney U or Kruskal-Wallis test as appropriate. Contingency tables were analyzed by Fisher's exact test. Correlation studies were performed by the Spearman non-parametric test. Differences were considered significant if P<0.05. Allele frequencies between the groups of case and control subjects were estimated by allele counting and tested for deviation from Hardy-Weinberg equilibrium (HWE) by the software program DeFinetti (https://ihg.gsf.de/cgi-bin/hw/hwa1.pl). For the significant deviation threshold, we used a P value of <0.05.
SNP data were analyzed using SPSS software version 22 (SPSS Inc., Chicago, IL, USA). Logistic regression analyses adjusted for age and gender were used to evaluate the association between YAP1 genotypes and asthma, its intermediate phenotypes, the discretized (normal/high) serum IgE and discretized (normal/high) eosinophil levels (see at Subjects) or different phenotypes. Additionally, multinomial logistic regression adjusted for age and gender was used for the analysis of YAP1 SNPs and GINA statuses. Confidence intervals (CIs) were calculated at the 95% level. Multiple comparisons were corrected for using the Benjamini-Hochberg correction, and a new significance level of P=0.004 with the false discovery rate (FDR) <6.5% was estimated. Haplotype analysis was carried out with the Haploview 4.2 program (Broad Institute of MIT, Cambridge, MA, USA). Odds ratios (ORs) for haplotypes were counted by VassarStats software (http://vassarstats.net/index.html).
Bayesian statistical analysis
Earlier, we have developed an alternative, a systems biological statistical method, named Bayesian network based Bayesian multilevel analysis of relevance (BN-BMLA). Bayesian networks offer a rich language for genetic association studies, because they exhaustively and exactly represent strongly relevant variables and their interactions through the Markov Blanket Set and Markov Blanket Graph features and they are able to evaluate multiple targets. Furthermore, this Bayesian global relevance analysis method provides posterior probabilities, which are direct statements about hypotheses; thus, it can also be used to construct probabilistic data analytic knowledge bases in genetic association studies to support complex querying, off-line meta-analysis, and fusion with background knowledge.18192021
We have previously described the BN-BMLA method in detail,15222324 so that the following only briefly summarizes this approach.
A Bayesian network is a directed acyclic graph (DAG) that aids the discovery of various dependency relations between random variables by representing their joint probability distribution. A node in the network represents a variable, and edges connecting 2 nodes represent direct dependency between those variables. To find the dependence relations of the variables, a DAG that best describes the dataset must be found. In most cases, there are many DAGs with non-negligible posteriors, but certain structural features may be extracted accurately. Such a feature is based on the concept of strong relevance of a single variable or a set of variables. Bayesian learning allows the evaluation of the strength of the data indicating the presence of a certain feature by evaluating its a posteriori probability.
The a posteriori probability can be calculated for strongly relevant variable sets with regard to a target variable. The strongly relevant variables have direct impacts on the target. The a posteriori probability of the strong relevance is between 0 and 1, where 1 means that the target (e.g., phenotypes of asthma) most certainly has a dependency relationship with a predictor (e.g., SNP), on the other hand 0 means that there is no such relationship. Posterior probabilities of strong relevance greater than or equal to 0.5 are regarded as relevant, and above 0.75 as convincing.18192021
In the present study, 29 SNP in the YAP1, FRMD6, and BIRC5 genes (genotyped with the same methods and on the same populations;1516
Supplementary Table S1) and the characteristics of the patients detailed in Table 2 were involved in the BN-BMLA analysis.
RESULTS
Gene expression in induced sputum
In the induced sputum of 18 asthmatic patients and 10 control subjects, we measured the gene expression level of 7 members of the Hippo/YAP1 pathway. The expression of all genes was detected in both cases and controls. The mean gene expression level of YAP1 was slightly lower in the asthmatic patients than in the control subjects (P=0.032, Fig. 2A). No other differences were detected in this respect. We investigated whether within the asthma group there were differences in gene expression between the subgroups of patients defined by GINA status, but no significant differences were found.
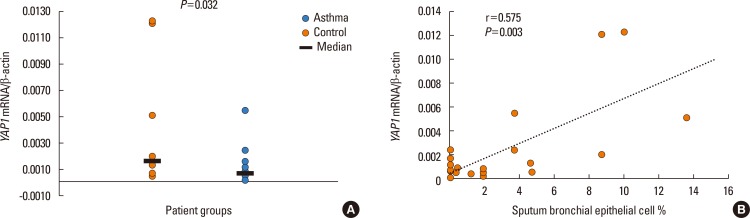
Fig. 2
(A) YAP1 mRNA levels in the induced sputum of asthmatic patients (n=18) and controls (n=10). (B) Relationship between YAP1 mRNA and bronchial epithelial cell levels. The mRNA levels were normalized and expressed according to the delta delta Ct method. The Mann-Whitney U test (A) and Spearmann's non-parametric correlation (B) were used.
![]()
During the correlation studies, we found a significant and positive correlation between YAP1 mRNA levels and sputum bronchial epithelial cells (r=0.575, P=0.003; Fig. 2B). There was a significant and negative correlation between TAZ mRNA and sputum neutrophils (r=−0.509, P=0.009), and STK4 showed a significant and positive correlation with sputum eosinophils (r=0.425, P=0.034). There was no significant correlation of YAP1, TAZ, LATS1, LATS2, SAV1, STK3, or STK4 gene expression with other cellular components, asthma severity, age, gender, airway inflammation, or ICS dose.
SNP association study results
We examined whether any of the SNPs in the YAP1 gene influence the susceptibility of asthma or different phenotypes. The statistically significant genotyping results are summarized in Table 4. There was no significant association of the SNPs with asthma susceptibility, allergic status, inhalative, outdoor, indoor allergies, allergic and non-allergic asthma, comorbidities of rhinitis and conjunctivitis or serum IgE and eosinophil levels. However, SNP rs2846836 was significantly associated with exercise-induced asthma (OR=2.1 [1.3-3.4], P=0.004, power=0.83; Table 4 and Supplementary Fig. S1A). Additionally, the distribution of genotypes of SNP rs11225138 showed a significant difference between GINA 1-2 and GINA 3-4 statuses in a dominant model (OR=2.8 [1.4-5.6], P=0.003, power=0.83; Table 4 and Supplementary Fig. S1B).
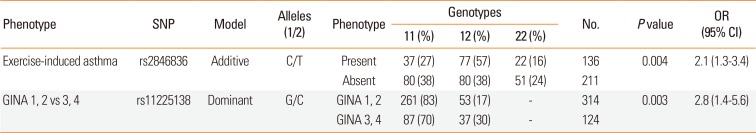
Table 4
Significant results of the association analysis of YAP1 SNPs with asthma phenotypes
![]()
In order to find more evidence for the associations, we also conducted haplotype analyses. We found a significant difference between patients of GINA 2 and GINA 3 when we compared the frequencies of a haplotype formed by the rare alleles of SNPs rs1426398 and rs11225138, where the frequency of TC haplotype was more prevalent in GINA 3 than in GINA 2 (28% vs 8%, P=10−7). Furthermore, the CA haplotype from SNPs rs11225138 and rs1426394, also showed a significant difference when patients with GINA 3 were compared to GINA 2 asthmatics (26% vs. 7%, P=10−7). When we included more than 2 SNPs in the analysis, additional associations were found. Corresponding results are shown in Supplementary Table S2 and Supplementary Fig. S2.
Western blot analysis of induced sputum
Western blots were carried out on the proteins of the Hippo/YAP1 pathway whose genetic variations showed significant associations with asthma or phenotypes. Earlier, we found a strong significant association between a genetic variation in the FRMD6 gene and asthma (P<0.001, Supplementary Table S1),15 and in the present study genetic variations and haplotypes in the YAP1 gene was associated with different phenotypes of asthma.
The signal for the FRMD6 protein could be detected in all sputum samples from both asthmatic patients and control subjects. Interestingly, although the YAP1 protein was not detected in the sputum samples of the healthy controls, it was seen in the sputum samples of the mild asthmatics (GINA 1, 2) and was absent in the sputum of severe (GINA 3, 4) asthmatics (Supplementary Fig. S3).
BN-BMLA
Based on the genotyping results of 29 SNPs in YAP1, FRMD6, and BIRC5 genes, the laboratory data and characteristics of the asthmatic patients detailed in Table 2, the posterior probabilities of relevance between the variables with respect to target variables were calculated by BN-BMLA.
Table 5 shows the most relevant variables with high posterior probabilities according to the BN-BMLA analysis. As expected, e.g., IgE levels or inhalative allergy was highly relevant to allergic asthma or eosinophil levels and allergic conjunctivitis to allergic rhinitis.
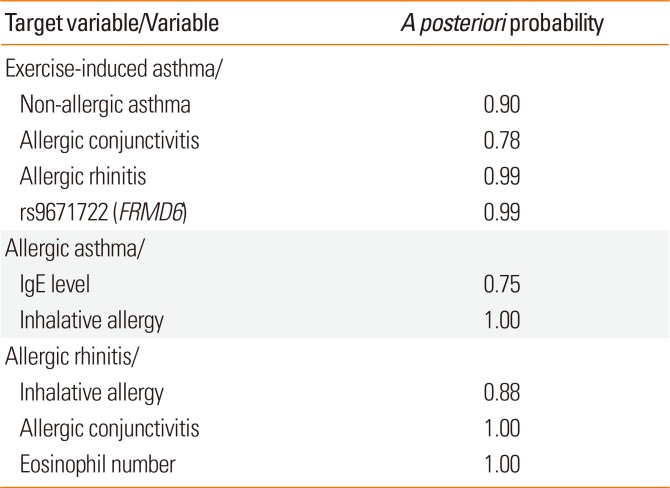
Table 5
The most relevant results of the BN-BMLA statistical method
![]()
In the case of genetic variations, no direct SNP-SNP or gene-gene interactions were found. The most relevant association was between rs9671722 in the FRMD6 gene and exercise-induced asthma with a posteriori probability of strong relevance of 0.99. Fig. 3 shows the most likely subgraph of the dependence structure of the variables. This structure suggests a direct relevance of rs9671722 to exercise-induced asthma, while another SNP (rs3751464) of the FRMD6 gene was found to be directly relevant to allergic rhinitis and transitively associated through allergic rhinitis with exercise-induced asthma.
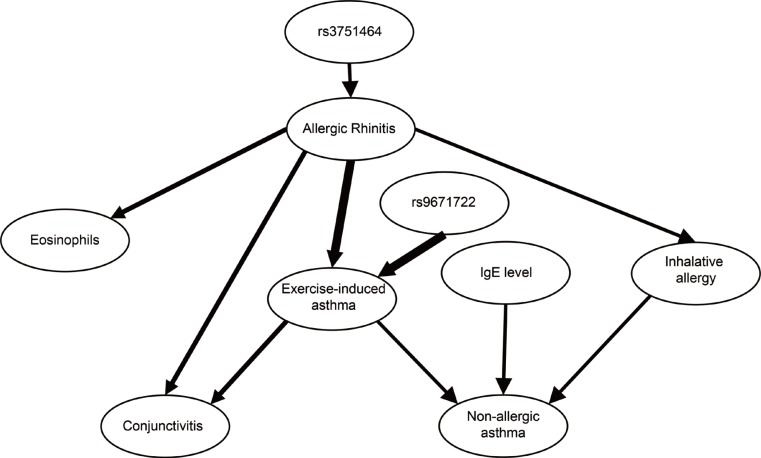
The relationship was confirmed by logistic regression which showed that patients with GG genotypes of the rs9671722 SNP had the highest odds of having both allergic rhinitis and exercise-induced asthma (OR=18.0 [5.9–54.9], P=3.7E-7; Supplementary Table S3). The interaction term in the logistic regression model was significant (P=0.010).
In Supplementary Fig. S4, another graph is presented which shows a dendrogram of the subsets of strongly relevant variables with respect to allergic rhinitis.
DISCUSSION
Recent studies have shown that inflammation caused by tissue damage or microbial invasion not only plays an important role in host defense, but also triggers regeneration and repair. It was also shown that house dust mite-induced asthma leads to a significant increase in reactive oxygen species (ROS) production and DNA damage in lung tissues, especially in the bronchial epithelium.25 However, the mechanisms, by which inflammation, ROS and DNA damage stimulate regenerative responses remain unclear.
Originally, it was found that the Hippo/YAP1 pathway played an important role in the regulation of organ size, but recent studies have indicated that YAP1 protein could also be detected in peripheral respiratory epithelial cells of the adult mouse lung.14 Furthermore, it was shown, that after depletion of club cells in the lungs, the distribution and intensity of YAP1 staining were increased. After 10 days, when the regeneration of the bronchiolar epithelium was complete, the YAP1 level and distribution was similar to that in the uninjured airway.14 In addition, multiple lines of evidence link the Hippo pathway to oxidative stress or ROS-initiated signaling pathways and various pathological processes. YAP1, the major Hippo downstream target, also mediates ROS-triggered signaling.26 These findings in mice suggest that the Hippo/YAP1 pathway can also play an important role in the regeneration processes in human asthma.
Earlier, we carried out a partial genome screening in pediatric asthma and identified the FRMD6 gene which showed the most consistent association with asthma susceptibility and its role in asthma development was also confirmed in an animal model.15 In an independent study, we identified BIRC5, of which genetic variations influenced asthma susceptibility and expression changed significantly during asthma both in animal and human studies.16 As FRMD6 is a possible upstream mediator of the Hippo pathway and BIRC5 is a target gene of YAP1, these findings also indicated that the Hippo/YAP1 pathway might play a role in asthma.
In the present and earlier studies, we found that all the important members of the FRMD6/Hippo/YAP1 pathway were expressed in the human induced sputum in both asthmatic patients and healthy controls. The gene expression level of different components of the Hippo pathway correlated with diverse cell types. This may indicate the main sources of these mRNA in the sputum, suggesting that the regulation of their expressions may be different in these cells and that the genes have additional, diverse functions in these cells.
The YAP1 gene expression level was correlated with the number of sputum bronchial epithelial cells, suggesting its possible origin. The YAP1 mRNA level was not correlated to the severity of asthma or other asthma phenotypes. On Western blot, however, the protein product for YAP1 was detected only in mild asthmatics and was not seen in controls or in severe asthmatics.
It has been suggested that YAP1 is regulated by the Hippo pathway which is a kinase cascade that ends up phosphorylating and inhibiting the protein. Recently, several studies have indicated that Hippo-mediated YAP1 phosphorylation is really a fundamental input for YAP1 regulation, but not the only one. Inflammation, DNA damage, and ROS or mechanical signals can represent separate signals with partly independent pathways that can regulate YAP1 phosphorylation and activity.2526272829
The mechanisms of YAP1 inhibition by phosphorylation are nuclear exclusion, sequestration in the cytoplasm, or proteasomal degradation. In the present study, YAP1 mRNA was detected in all samples, but YAP1 protein was detected only in mild asthmatics. Although the detection level of the RT-PCR is lower than that of the Western blot, this finding is in agreement with the previous notion that YAP1 activity is also and perhaps mainly regulated on the protein level. Because of the low number of patients and the detection method, this can only be regarded as a preliminary finding, but based on the above-described observations, the appearance of YAP1 in the mild asthmatics can be explained by the asthma-associated tissue damage that induces regeneration where the Hippo/YAP1 pathway can play an important role.303132 Inflammation, DNA damage, and elevated ROS in the airway epithelium were all found to be associated with an increased YAP1 protein level.26 Presently, it cannot be explained why the YAP1 protein could not be detected in the sputum samples of severe asthmatics. However, it can be hypothesized that by an unknown mechanism, YAP1 is less or not activated in the lung of severe asthmatics which can result in a defected regeneration process in the airways which can contribute to irreversible organ damage and the severity of symptoms in these patients.
Here, we have to mention some limitations of our study. First, although Western blot can determine the molecular weight of protein and in this way has a higher specificity compared to e.g., enzyme-linked immunosorbent assay (ELISA), it is less sensitive to and less capable of quantitative measurement. Secondly, in this study we did not differentiate between dephosphorylated YAP1 and phosphorylated YAP1. Also, as we have no available data on when ICS were administered, we cannot exclude the possibility that various time intervals between ICS administration and sputum induction may influence our results.
The possible role of the Hippo/YAP1 pathway in asthma is further supported by the results that a genetic variation in the YAP1 gene was found to be associated with exercise-induced asthma and that distributions of certain haplotypes differed significantly between different asthma GINA statuses. This latter observation, in line with the lack of YAP1 protein in the induced sputum of severe asthmatics, also supports the finding that haplotypes in the YAP1 gene were associated with the severity of the disease. It must be noted, however, that there can be differences between childhood and adult asthma; thus, the genetic associations must be confirmed in a well characterized adult population.
Based on our genotyping results and the characteristics of the asthmatic patients, we searched for the most probable interaction networks with respect to different target variables. We also wanted to know whether there would be interactions between the three genes (FRMD6, YAP1, and BIRC5) whose genetic variations could be associated with asthma or asthma phenotypes in this population. Using the BN-BMLA method, no interaction was found between these genes, indicating that the genetic variations in the 3 genes influenced disease susceptibility independently from one another and that the studied population was too small to detect such interactions.
Within the asthma group, 2 genetic variations in the FRMD6 gene proved to be the most relevant to exercise-induced asthma and allergic rhinitis. The 2 SNPs through allergic rhinitis and exercise-induced asthma were in epistatic interaction with each other. The term exercise-induced asthma describes transient narrowing of the airways after exercise. Presently, the exact mechanism of exercise-induced asthma is not known, but as mouth breathing is common during exercise, there is increased penetration of pollutants, cold air, and allergens into the airways that can lead to epithelial damage, inflammation, and remodeling.3334 Based on the literature and our findings, it is not possible to explain the relationship between the variations in the FRMD6 gene, rhinitis, and exercise-induced asthma, but a possible hypothesis can be that variations in the gene can weaken the regeneration capacity of the Hippo pathway which can lead to persistent epithelial damage and asthma symptoms in genetically susceptible individuals.
CONCLUSION
This study provides additional evidence that the FRMD6/Hippo/YAP1 pathways might play a role in the pathogenesis of asthma and its different subtypes. Naturally, genetic associations must be confirmed in independent populations and it would be also interesting to reveal how exactly the activity of YAP1 protein is regulated in the airways of asthmatic patients. If additional studies can confirm that YAP1-associated pathways play a role in the regeneration processes in airway inflammations, these pathways can be potential novel therapeutic targets in asthma and other inflammatory airway diseases.
 XML Download
XML Download