

 PDF
PDF Citation
Citation Print
Print



BACKGROUND
Ovarian cancer is most deadly gynecologic malignancies worldwide. There were an estimated 239,000 new cases with 152,000 deaths in 2012 [1]. This high mortality is mainly due to late diagnosis with less than 20% of ovarian cancer patients are diagnosed at an early stage (International Federation of Gynecology and Obstetrics [FIGO] stage I and II). The 5-year survival rate of these patients is over 90%. However, this number is less than 30% in patients in an advanced stage (FIGO stage III and IV) as therapies become increasingly ineffective in treating metastatic ovarian cancer [2].
Most ovarian cancer patients undergo debulking surgery followed by chemotherapy. While approximately 75% of patients initially respond to the platinum/paclitaxel-based chemotherapy, most of them relapse with chemoresistance which results in treatment failure and causes over 90% of deaths [3]. Thus, understanding the molecular mechanisms underlying this drug resistance is important for the development of effective therapies to improve ovarian cancer patients' outcome.
OVARIAN CANCER STEM/TUMOR-INITIATING CELLS
Cancer stem cells (CSCs) is a subpopulation of tumor cells with self-renewal and differentiation properties that can sustain tumor growth and recapitulate a heterogeneous tumor [4]. CSCs have been identified in hematologic malignancies and various solid tumors [56789]. Experimental evidence for the existence of ovarian CSCs was first reported in 2005, when Bapat and colleagues [10] identified a tumorigenic clone from malignant ascites of a patient with ovarian cancer through multilayer spheroid culture. Thereafter, ovarian CSCs have been isolated in clinical specimens by various methods based on phenotypic and functional properties of CSC, such as the capability to form tumor spheroids under suspension culture, the efflux capacity of fluorescent dye Hoechst 33258 and stem cell marker expression [111213].
CD44 is a cell-surface glycoprotein of hyaluronate receptor that plays a role in tumor stemness, recurrence and drug resistance in ovarian cancer. Paik et al. [14] identified CD44 as a marker for fallopian tube epithelial stem-like cells (FTESCs), and further suggested a role of FTESC in the initiation of serous tumors. CD44 in combination with other markers, such as CD117 [15], MyD88 [16], and CD24 [17] have been extensively used for ovarian CSCs isolation. For instance, Zhang et al. [15] isolated CD44+/CD117+ ovarian CSCs that were fully capable of re-generating the original tumor phenotype in mice, and were found to exhibit greater resistance (3.1–16.1 folds) to cisplatin and paclitaxel as compared to cells cultured under differentiating conditions. Several recent studies have shown that CD44 overexpression in ovarian cancer is associated with poor prognosis [181920]. Specifically, Gao et al. [20] reported higher expression of CD44 in metastatic/recurrent ovarian cancer tissue samples as compared with matched primary tumor samples, and there is a significant association between CD44 expression and unfavorable prognosis. Further, knocking down of CD44 increased tumor cells' sensitivity to paclitaxel, indicating that CD44 up-regulation might be a critical event in the development of drug resistance in ovarian cancer [20].
CD133 (prominin-1) is a pentaspan transmembrane protein initially recognized as a marker for human hematopoietic stem cells [21]. CD133 has been defined as a CSC marker in various tumors, including ovarian cancer. It has been documented that CD133+ ovarian cancer cells possessed tumorigenic and aggressive capacity, as well as enhanced resistance to chemotherapies compared with CD133− cells [222324]. In particular, Baba and colleagues [24] reported the IC50 value of cisplatin for CD133+ epithelial ovarian cancer cells was higher than that for CD133− cells, indicating a greater chemoresistance in CD133+ cells. They further demonstrated that mRNA expression of CD133 correlates with chemoresistance capability of CD133+ cells. Similarly, lower cisplatin sensitivity and higher breast cancer resistance protein (ATP-binding cassette sub-family G member 2 [ABCG2]) gene expression which is implicated in drug efflux were detected in C-X-C chemokine receptor type 4 (CXCR4)+CD133+ CSCs in comparison with non-CSCs counterparts, suggesting a chemoresistant phenotype in CXCR4+CD133+ ovarian CSCs [25]. Moreover, CD133 overexpression is associated with ovarian cancer patients' response to treatment and clinical outcome. Zhang et al. [26] reported a correlation of CD133 expression with high-grade ovarian serous carcinoma, advanced stage disease, ascites levels, lack of response to chemotherapy, shorter overall survival time, and reduced disease-free survival. Interestingly, compared with matched primary tumors, the percentage of CD133+ cells in recurrent tumors is increased from 6.3% to 34.5% in patients with platinum-resistant recurrence [27]. These studies support a clinical significance of CD133 in ovarian cancer chemoresistance.
CD117, also known as stem cell growth factor receptor or c-Kit, is a tyrosine kinase oncoprotein. CD117+ ovarian cancer cells isolated from human ovarian serous carcinoma tissues and ascites exhibited 100-fold higher tumorigenic capability than CD117− cells, and as few as 103 CD117+ cells were sufficient to self-renew and give rise to heterogeneous tumors in mice [28]. Besides, it was further found CD117 expression was statistically correlated with resistance to platinum-paclitaxel chemotherapy, as only 10% (1/10) CD117+ patients were sensitive to chemotherapy while a sensitivity was found in 60% (9/15) CD117− group, and correlated with shorter recurrence intervals [28]. These results indicate that CD117+ CSCs may escape from conventional chemotherapy, possess the ability to regenerate tumors and cause relapse after therapy. Based on an analysis of 56 patients with advanced serous ovarian cancer, the expression of CD117 was significantly correlated with worse response to first-line chemotherapy, decreased treatment-free interval and shorter overall survival [29], further supporting the importance of CD117 in ovarian cancer chemoresistance. Moreover, our group demonstrated that ovarian CSCs that showed high CD117 expression exhibited chemoresistance to clinically relevant doses of cisplatin and paclitaxel [30]. We further illustrated that hypoxia or HIF-1α could upregulate the expression of CD117 which promotes chemoresistance through a Wnt/β-catenin pathway. Knockdown or inhibition of CD117 increased chemosensitivity both in vitro and in vivo [30]. These results support the clinical relevance of CD117 in stemness and chemoresistance.
The aldehyde dehydrogenase (ALDH) superfamily comprises 19 isozymes that catalyze the oxidation of aldehyde. ALDH1A1, a major isoform in CSCs, has been described as a candidate ovarian CSC marker and is associated with chemoresistance [3132]. Landen and colleagues first identified ALDH1A1+ cells possessing CSC phenotype in ovarian cancer cell lines. High ALDH expression was significantly associated with poor progression-free survival in ovarian cancer patients, and knockdown of ALDH1A1 sensitized ALDH1A1+ cells to taxane and platinum [32]. Silva et al. [31] found that the percentage of ALDH+ in SKOV-3 cells increased with an increase in the cisplatin dose, and these isolated ALDH+ showed greater viability and faster recovery after cisplatin treatment than ALDH− cells, indicating greater chemoresistance in ALDH+ than ALDH− cells. They further isolated ALDH+CD133+ cells that were able to initiate xenograft tumors with only 11 cells and found that the presence of ALDH+CD133+ cells in primary ovarian tumors predicts worse outcome in patients [31]. These findings indicate a role for ALDH1+ CSCs in drug resistance and may be a chemotherapy target for ovarian cancer.
CD24 and epithelial cell adhesion molecule (EpCAM) are putative ovarian CSCs markers, which also play a role in chemoresistance. CD24 is a glycosylphosphatidylinositol-linked cell surface adhesion molecule that is frequently detected in invasive ovarian carcinomas while usually absent in normal ovarian surface epithelium and adenomas [33]. High cytoplasmic CD24 expression in the invasive ovarian carcinomas predicts shorter overall survival (37 months vs. 98 months) [33]. CD24+ cells isolated from human ovarian tumors were relatively more resistant to cisplatin than the CD24− counterpart [34]. Donahoe group isolated CD44+CD24+EpCAM+ E-cad− subpopulation that possesses CSC characteristics with enhanced tumorigenic capacity in vivo and chemoresistance to doxorubicin, cisplatin, and paclitaxel therapeutics [35]. These CSCs could be stimulated by doxorubicin and preferentially inhibited by Müllerian inhibiting substance [35]. Tayama et al. [36] found an elevated expression of EpCAM in ovarian tumor tissues in patients after receiving platinum-based chemotherapy compared with that in corresponding tissues before chemotherapy, and patients with high EpCAM expression are less likely to respond to first-line platinum-based chemotherapy and are associated with poor survival [36].
The mechanisms underlying chemoresistance of CSCs are unclear. Drug efflux through ATP-binding cassette (ABC) transporters has been described as one of the reasons (Fig. 1). The up-regulation of 2 ABC transporters, P-glycoprotein (P-gp or ABCB1) and ABCG2, were frequently observed in ovarian CSCs [1230]. The quiescent state of CSCs, which is essential for preserving the self-renewal function of stem cells, is also a critical factor in the drug resistance. Unlike the rapidly dividing cells, the relatively quiescent CSCs are insensitive to most of the chemotherapeutic drugs [37]. Indeed, quiescent CSCs that display high chemoresistant capability have been identified in ovarian cancer cells lines [38] and tumor tissue specimens [34]. In addition, DNA repair and autophagy are also implicated in the chemoresistance of ovarian CSCs. For example, polymerase η (Pol η) enhanced cisplatin-induced apoptosis and blocked the cisplatin-induced enrichment of ovarian CSCs, suggesting that an increase in expression of DNA Pol η in CD44+/CD117+ contributes to drug resistance [39]. Similarly, CD44+/CD117+ ovarian CSCs have higher autophagy levels than non-CSCs counterparts, and inhibition of autophagy impairs the ability of ovarian CSCs to resist chemotherapy in vitro and in vivo [40].
 | Fig. 1Schematic representation of CSCs in ovarian cancer chemoresistance. Chemotherapeutics fail to eliminate CSCs which can regenerate the entire tumor and ultimately result in a relapse. Ovarian CSCs are resistant to chemotherapy due to several mechanisms including increased drug efflux, quiescence, enhanced DNA repair, autophagy, etc.
ALDH, aldehyde dehydrogenase; CSC, cancer stem cell; EpCAM, epithelial cell adhesion molecule.
|
OVARIAN CANCER METABOLISM
Metabolic reprogramming was first seen as a consequence of oncogene activation. Later, loss-of-function mutations of metabolic enzymes have also been observed in the pathogenesis of several hereditary forms of cancer such as ovarian cancer and breast cancer. These data also suggest that metabolic reprogramming could be the cause, rather than the consequence, of tumor transformation [41]. Glycolysis is a coordinated series of chemical reactions that convert one molecule of glucose into lactate, and generate 2 molecules of ATP [42], which are the major source of energy of cancer cells [43]. Unlike normal tissues, tumor cells preferentially metabolize glucose by glycolysis instead of oxidative phosphorylation (OXPHOS), even in the presence of oxygen. The so called “Warburg effect” is one of the key steps of cancer development [4445]. This up-regulation of glycolysis will generate more lactate via lactate dehydrogenase. The acidity in the microenvironment is known to facilitate tumor invasion and metastasis [4647]. Furthermore, the accelerated glycolysis in tumors also contributes to chemoresistance [4849505152]. While the underlying mechanisms of action remain unclear, it has been shown that downregulation of glycolytic enzymes could improve or even overcome drug resistance [5354].
Many glycolytic inhibitors are currently under preclinical or clinical development (Fig. 2). Hexokinase 2 (HK2) is a key glycolytic enzyme that phosphorylates glucose into glucose-6-phosphate (G6P). In ovarian cancer, HK2 is highly expressed in epithelial ovarian cancer tissues, especially the serous subtype, but is scanty in normal, benign, and borderline ovarian tumors. Also, HK2 expression is strongly linked to chemoresistance [5556]. Collectively, HK2 may be a promising target for ovarian cancer treatment. Lonidamine, a HK2 inhibitor, has been used for cancer treatment in clinics in Europe [57]. Lonidamine has been shown to revert the resistance to cisplatin and paclitaxel in advanced ovarian cancer [58].
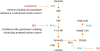 | Fig. 2Schematic of key glycolysis and lipogenesis enzymes and inhibitors.
3PO, 3-(3-pyridinyl)-1-(4-pyridinyl)-2-propen-1-one; DIC, dicumarol; F-1,6-BP, fructose 1,6-bisphosphate; F6P, fructose-6-phosphate; FASN, fatty acid synthase; G6P, glucose-6-phosphate; HK2, hexokinase 2; LDH, lactate dehydrogenase; PDH, phosphate dehydrogenase; PDK1, 3-phosphoinositide-dependent protein kinase-1; PFK-1, phosphofructokinase-1.
|
Phosphofructokinase-1 (PFK-1) catalyzes the conversion of fructose 6-phosphate and ATP to fructose 1,6-bisphosphate and ADP. PFK-1 regulates the highly exergonic and irreversible step of glycolysis regardless of the cancer type. It has been demonstrated that loss of 6-phosphofructo-2-kinase/fructose-2, 6-bisphosphatase 3/4 (PFKFB3/4), the key enzyme regulating PFK-1 activity, can increase caspase 3/7 activity and reactive oxygen species (ROS) levels, thus enhancing mitotic cell death via phosphoinositide 3-kinase (PI3K)/Akt pathway when combined with paclitaxel treatment in mitotically arrested ovarian cancer [59]. The 3-(3-pyridinyl)-1-(4-pyridinyl)-2-propen-1-one (3PO) is an inhibitor of PFKFB3, which has been demonstrated to suppress glycolysis and tumor growth [60].
The 3-phosphoinositide-dependent protein kinase-1 (PDK1) is a kinase enzyme which acts to inactivate the enzyme pyruvate dehydrogenase by phosphorylation, resulting in the suppression of OXPHOS in mitochondria [48]. PDK1 is almost absent in normal ovaries, and 60% of benign ovarian tumors. Overexpression of PDK1 was evident in borderline and low- to high-grade ovarian tumors and correlates with survival in high-grade tumors [6162]. PDK1 is also highly expressed in chemoresistant ovarian cancer cells [63]. Dicumarol could inhibit PDK1 activity, shifting the glucose metabolism from aerobic glycolysis to OXPHOS, increasing ROS levels, and inducing apoptosis in vitro and in vivo in ovarian cancer [64].
Lipid is strongly associated with ovarian cancer progression. Several specific lipids are increased in ovarian cancer patients [65]. Fatty acid synthase (FASN) is one of the most important biosynthetic enzymes in lipogenesis. It has been reported that FASN expression significantly correlates with tumor grade and FIGO staging. Patients with FASN overexpression tend to have a worse overall survival rate [66]. Ovarian tumors overexpressed FASN can be inhibited by FASN blockers, whereas normal tissues are FASN negative [67]. ERBB-PI3K-mechanistic target of rapamycin complex 1 (mTORC1) up-regulates FASN through the activation of the transcription factor sterol regulatory element-binding protein (SREBP)-1c [68]. FASN in turn can stimulate PI3K-mTORC1 signaling [6970]. Furthermore, inhibition of FASN can induce apoptosis in platinum-resistant ovarian cancer cells and may reverse cisplatin resistance [71].
Epidemiological and clinical studies have suggested that polyunsaturated fatty acids (PUFAs) could have health benefits and therapeutic effects in cancer treatment. A recent study demonstrated that ω-3 and ω-6 PUFAs could induce cell death in ovarian cancer cell lines through ROS-dependent MAP kinase activation [72]. Moreover, Li et al. [73] showed that ovarian CSCs identified as ALDH+/CD133+ population have higher levels of unsaturated fatty acids (UFAs) and stearoyl-CoA desaturase-1 (SCD1, an enzyme produces mono-UFAs) compared to non-CSCs (ALDH−/CD133−). Knockdown or inhibition of SCD1 could reduce CSC markers ALDH1A1, Sox2, Nanog, and Oct-4 expression in ovarian CSCs and patients derived CSCs. Furthermore, inhibition of lipid desaturation reduces the nuclear factor (NF)-κB activity in ovarian CSCs [73].
CSC METABOLISM AND CHEMORESISTANCE
Several studies have indicated distinct metabolic features exist in CSCs. Some studies have shown that CSCs bear more active glycolytic activity than differentiated tumor cells [74757677]. For example, by comparing metabolite profiles of ovarian CSC-like spheroids and parental cells, it is found that spheroid cells mainly relied on anaerobic glycolysis and involved direct glucose oxidation in the pentose cycle [78]. Similarly, more active glycolysis was observed in CSCs isolated in rodent ovarian surface epithelium compared with parental cells [79]. This distinct glycolytic metabolism profile in CSCs indicates that CSCs are endowed with the capacity to survive under stress conditions, in particular hypoxia, and may contribute to chemoresistance [78]. In the contrary, CD44+CD117+ ovarian CSCs showed an elevated expression of enzymes associated with OXPHOS with higher mitochondrial ROS production, suggesting that the mitochondria electron respiratory chain is preferentially utilized [80]. Mitochondrial metabolism is also described in lung [81] and pancreatic CSCs, and that this OXPHOS-dependent metabolic phenotype seems to be associated with a selective advantage to survive in nutrient starvation and other environmental stress conditions [808283]. It is still controversial regarding if CSCs use glycolytic or mitochondrial respiration pathway in comparison with non-CSCs [778485]. However, it is noted that the metabolic plasticity that CSCs may alter metabolic phenotypes in adaptation to microenvironmental fluctuations [8386]. Moreover, elevated fatty acid β-oxidation (FAO) activity was found in ovarian CSCs in comparison with non-CSC counterpart [80]. In breast cancer, an elevated FAO is responsible for self-renewal and chemoresistance of the CSCs [87]. These biochemical characteristics suggest a possibility that the metabolism in CSCs may be vulnerable to therapeutic intervention.
 XML Download
XML Download