

 PDF
PDF Citation
Citation Print
Print



INTRODUCION
Ovarian cancer is a leading cause of mortality from gynecological malignancy. It is also the second most common gynecological cancer and one of the five leading causes of cancer-related death amongst females [1]. Most ovarian cancer patients respond to the standard therapy of initial cytoreductive surgery followed by taxane- and platinum-based chemotherapy; however, 70% of patients experience a relapse of the cancer within 2 years [23]. Recurrent cancer is typically less responsive to chemotherapy regimens, possibly because the cancer cells develop resistance mechanisms to the drugs through changes in their genetic and acquired natures [4]. The development of chemosensitization strategies that can overcome chemoresistance is an important area of research.
Emerging evidence suggests that cancers contain a subpopulation of stem-like cells, called cancer stem cells (CSCs), with self-renewing abilities, high chemoresistance, and aggressive characteristics. CSCs are believed to contribute to tumor heterogeneity and to play a significant role in treatment failure and tumor relapse [5]. Under appropriate culture conditions, individual CSCs are capable of growing into 3-dimensional structures called spheroids or tumorspheres. Spheroid-forming cells are characterized by the upregulation of stem cell genes, self-renewal, and high proliferative and differentiation potentials. Furthermore, spheroid-forming cells are more aggressive in growth, migration, invasion, scratch recovery, clonogenic survival, and anchorage-independent growth, and are more resistant to chemotherapy in vitro [6]. Bapat et al. reported the identification of cells with the characteristics of cancer stem-like cells in ovarian malignancy [7].
Recent research has established that expression of CD44, a surface transmembrane glycoprotein, is associated with “stemness” in ovarian cancer cells (OCCs). CD44 is involved in cell-cell and cell-matrix interactions that affect cellular growth, differentiation, and motility. Increased expression of CD44 correlates with resistance to chemotherapy and poor prognoses [8]. Expression of CD117, a receptor tyrosine kinase also known as c-kit, is also associated with potential existence of ovarian cancer stem-like cells with properties of self-renewal, differentiation, a high tumorigenic potential, and chemoresistance [9]. Therapies targeting leukemia, breast, prostate, and colorectal CSCs have shown antitumor activities and inhibition of cancer cell proliferation [10]. The present study aims to further develop ovarian cancer treatment by targeting ovarian cancer stem cells (OCSCs) marked by CD44 and CD117 expression.
Salinomycin is a monocarboxylic polyether antibiotic isolated from Streptomyces albus and originally used as an agricultural antimicrobial agent. Gupta et al. [11] first reported salinomycin as an effective agent against breast CSCs in 2009. Current research shows that salinomycin can overcome drug resistance in human cancer and suppress cancer cell and CSC proliferation by inducing apoptosis [12]. Salinomycin has been found to inhibit breast CSC-induced tumorsphere formation, overcome adenosine triphosphate-binding cassette (ABC) transporter-mediated multidrug resistance and apoptosis resistance in human acute myeloid leukemia stem cells, and target CSCs of hepatocellular carcinoma and brain, pancreatic, lung, and prostate cancer [131415161718]. These data suggest that salinomycin can potentially be a CSC targeting agent in human cancer treatment. However, the mechanisms of the anticancer effects of salinomycin remain unclear.
In the present study, we conducted spheroid-forming and apoptosis assays to investigate whether salinomycin reduces stemness and induces apoptosis in human OCSCs. Our findings shed light on the potential effectiveness of treatment that combines salinomycin with taxane-based chemotherapy.
METERIALS AND METHODS
1. Primary culture conditions of OCCs
Ascites was collected from patients diagnosed with advanced stage high-grade serous ovarian carcinoma, after obtaining written informed consent under protocols approved by the Ethical Committee of the Keimyung University Dongsan Medical Center, Korea. Epithelial OCCs were isolated using standard procedures. The ascites fluid was centrifuged at room temperature for 5 minutes at 1,500 rpm. The cell pellets were filtered (100-μm cell strainer) and washed twice with phosphate buffered saline (PBS), and red blood cells (RBCs) were removed by Histopaque-1077 (Sigma-Aldrich, St. Louis, MO, USA). The single cells were dispersed in complete medium composed of DMEM/F12 with 100 units/mL antibiotics. The conditioned medium was changed every two days.
2. Isolation of CSCs
The CD44+CD117+ cells were sorted from the primary OCCs by using the magnetic-activated cell sorting (MACs; Miltenyi Biotec, Bergisch Gladbach, Germany). CD44+ subsets were isolated by using mouse antihuman CD44 antibody coupled to magnetic microbeads (Miltenyi Biotec), followed by the magnetic column selection or depletion. The resulting cells were depleted of CD117− subsets by using mouse antihuman CD117 antibody coupled to magnetic microbeads (Miltenyi Biotec). CD44+CD117+ cells were named as OCSCs.
3. Cell viability assay
The number of viable cell exposed to paclitaxel (PTX) and salinomycin was evaluated by a colorimetric 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay. Initially, cells were seeded in 96-well plate, and cultured for 24 hours to allow their adhesion to the plate. After pre-incubation, the culture medium was changed to experimental medium supplemented with dimethyl sulfoxide (DMSO, 0.1%) control, PTX (10–200 nM) and salinomycin (0.5–5.0 µM) alone or in combination for 48 hours. The intensity of the purple color formed by this assay is proportional to the number of viable cells. MTT reagent was added and incubated for an additional 4 hours at 37°C. The optical density (OD) was measured at 540 nm. The mean values and their standard deviations were calculated from triplicate experiments.
4. Spheroid forming assay
Tumor sphere culture was performed in ultra-low attachment 60 mm dishes (Corning, New York, NY, USA), serum-free DMEM/F12 medium, supplemented with 5 μg/mL insulin (Sigma-Aldrich), 20 ng/mL human recombinant epidermal growth factor (Invitrogen, Carlsbad, CA, USA), 10 ng/mL basic fibroblast growth factor (Invitrogen) and 0.4% bovine serum albumin (Sigma-Aldrich).
5. Protein isolation and immunoblotting
Cell (1×107/mL) extracts were prepared in 1×radioimmunoprecipitation assay (RIPA) buffer containing protease and phosphatase inhibitors with EDTA (Thermo Scientific, Rockford, IL, USA). Protein concentrations were measured using Protein Assay Reagent (Bio-Rad, Hercules, CA, USA) following the manufacturer’s protocol. Aliquots of protein were separated by 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis and transferred to nitrocellulose membrane (Bio-Rad). The membranes were blocked with Tris-buffered saline containing 5% skim milk (Becton Dickinson and Company, Oxford, UK) and 0.2% Tween 20 (Amresco Inc., Solon, OH, USA). The following primary antibodies were used: sex determining region Y-box 2 (SOX2, 1:1,000; Cell Signaling, Danvers, MA, USA), octamer-binding transcription factor 3/4 (OCT3/4, 1:1,000; Cell Signaling), death receptor 5 (DR5, 1:1,000; Koma Biotech Inc., Seoul, Korea), Bcl2-associated X protein (Bax, 1:1,000), B cell lymphoma 2 (Bcl-2, 1:1,000; Biotechnology Inc., Santa Cruz, CA, USA), X-linked inhibitor of apoptosis protein (XIAP, 1:1,000; Becton Dickinson and Company), Survivin (1:1,000; Biotechnology Inc.), and β-actin (1:10,000; Sigma-Aldrich). After reaction with horseradish peroxidase conjugated secondary antibodies (Biotechnology Inc.) bands on the membranes were visualized by an enhanced chemiluminescence system (Thermo Scientific) following the manufacturer’s suggested procedure. The density of respective bands was analyzed by the chemi-Doc XRS imaging system (Bio-Rad).
6. siRNA gene silencing
Transfection of siRNA was performed using ON-TARGETplus (Dharmacon, Lafayette, CO, USA) kit, according to manufacturer’s instruction. SMARTpool sequenced siRNA targeting SOX2 or non-targeting siRNA scrambled control (negative control) was diluted to a working stock solution of 5 μM in RNase free water. Transient transfection was done using the DharmaFECT transfection reagents. After 24 hours of post transfection, cells were used to perform immunobloting and MTT assay.
7. Apoptotic assessment by annexin V/propidium iodide (PI) staining
Annexin V assay was used to detect apoptotic cells by staining cells with both annexin V-fluorescein isothiocyanate (FITC) and PI. Cells were incubated for 48 hours at 37°C with PTX (20 nM) and salinomycin (0.5 µM). After washing with cold PBS, cells were re-suspended in 1×binding buffer. Staining of apoptotic cells was performed using the annexin V-FITC/PI apoptosis assay kit (Becton Dickinson and Company) according to the manufacturer’s instructions. Determination of apoptotic cells was performed in a NovoCyte flow cytometer using NovoExpress 1.0.2 software (ACEA Biosciences, San Diego, CA, USA).
8. Caspase-3 activity test
After drug treatment, cells were centrifuged at 2,000 rpm for 5 minutes to harvest. Cells (2×106) were then lysed in 50 µL of lysis buffer on ice for 10 minutes and centrifuged at 14,000 rpm for 5 minutes at 4°C. Collected supernatant (50 µL) was added to an equal volume of 2×reaction/DTT Mix supplemented with DEVD-pNA (50 µM) and incubated at 37°C for 3 hours. Caspase-3 activity was measured as the absorbance at 405 nm of the cleaved substrate pNA followed by ApoAlert Caspase Colorimetric Assay kits User Manual (Clontech Laboratories, Mountain View, CA, USA).
9. DNA fragmentation assay
Fragmented nucleosomal DNA was quantified by Cell Death Detection enzyme-linked immunosorbent assay (ELISA) Plus kit (Roche Diagnostics, Mannheim, Germany) as described in the manufacture’s manual. Supernatant (20 µL) was used to detect apoptosis with microplate reader (Tecan, Salzburg, Austria) at 405 nm. Background values were subtracted and OD values representing nucleosomal DNA fragments in Hsp90 inhibitor treated samples were compared with the values obtained from control cells and expressed as percentage (%) of control.
RESULTS
1. Inhibitory effect of salinomycin on OCSC proliferation
The potential of salinomycin to inhibit the growth of OCCs and OCSCs was determined using a MTT assay. In comparison with the control, PTX reduced the viability of OCCs in a dose-dependent manner but only slightly reduced the viability of OCSCs (Fig. 1A). On the other hand, salinomycin reduced the viability of OCSCs in a dose-dependent manner, while only slightly affecting OCCs (Fig. 1B). PTX (20 nM) inhibited the growth of OCCs and OCSCs by 27% and 11%, respectively. Inhibition of growth of OCCs and OCSCs by PTX (20 nM) combined with salinomycin (0.5 µM) was 58% and 59%, respectively (Fig. 1C). These findings suggest that salinomycin and PTX could have a synergistic effect for treating ovarian cancer.
Fig. 1
Salinomycin inhibits cell proliferation in human OCSCs. (A) Proliferation of OCCs and OCSCs treated with PTX (10, 20, 50, and 100 nM) was observed for 48 hours. (B) Proliferation of OCCs and OCSCs treated with salinomycin (0.5, 1, and 5 µM) was observed for 48 hours. (C) Cells were treated with DMSO (control) or PTX (20 nM) and salinomycin (0.5 µM) alone or in combination for 48 hours. Cell viability was measured using the MTT Assay. The values are average of ±standard deviation.
DMSO, dimethyl sulfoxide; MTT, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide; OCCs, ovarian cancer cells; OCSCs, ovarian cancer stem cells; PTX, paclitaxel; Sal, salinomycin.
*p<0.05.

2. Role of salinomycin on stemness
We examined whether salinomycin reduced stemness-related gene and protein expression in OCSCs. After treatment with a combination of PTX and salinomycin, OCSCs down-regulated expression of stemness-related mRNA and protein (Fig. 2A, B) such as SOX2 and OCT3/4. To elucidate the effect of SOX2, we examined OCT3/4 expression following SOX2 siRNA transfection in OCSCs (Fig. 2C). SOX2 silencing sensitized the OCSCs to the anti-cancer effect of PTX (Fig. 2D). Taken together, these data demonstrate that salinomycin affects stemness genes, and that the targeting of SOX2 enhances the effect of PTX on OCSCs.
Fig. 2
Effect of PTX and salinomycin treatment on stemness marker of OCSCs. Cells were exposed to PTX (20 nM) and salinomycin (0.5 µM) alone or in combination for 48 hours. Analysis of SOX2 and OCT3/4 mRNA (A) and protein (B) expression levels in OCSCs. Effect of silencing SOX2 on OCT3/4 expression in OCSCs. Cells were transfected with SOX2 siRNA and control siRNA. (C) After 24 hours transfection, SOX2, and OCT3/4 proteins were analyzed by western blotting. (D) Cell viability was determined by MTT assay. The values are average of ±standard deviation.
GAPDH, glyceraldehyde 3-phosphate dehydrogenase; MTT, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide; OCSCs, ovarian cancer stem cells; OCT3/4, octamer-binding transcription factor 3/4; PTX, paclitaxel; Sal, salinomycin; SOX2, sex determining region Y-box 2.
*p<0.05.
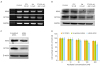
3. Effects of salinomycin on the spheroid-forming ability of OCSCs
Spheroid formation has been described as an important feature for the survival of OCSCs [17]. We evaluated the spheroid-forming abilities of OCSCs treated with PTX (20 nM), salinomycin (0.5 µM), a combination of both, and neither (as a control group). OCSCs were grown in nonadherent culture conditions in the absence or presence of salinomycin and PTX for 14 days, and spheroid formation was characterized in vitro (Fig. 3A). PTX-alone treatment reduced 46.6%, and salinomycin-alone treatment reduced 60.0% of the spheroid forming behavior. Cells treated with a combination of salinomycin and PTX showed significant reduction of 86.6% in spheroid formation compared with control cells (Fig. 3B).
Fig. 3
Salinomycin inhibits the formation of spheroids. (A) OCSCs were incubated with PTX and/or salinomycin for 14 days under spheroid forming conditions. The size of the spheroids was evaluated under a microscope. (B) Number of spheroid formation of OCSCs was counted. Scale bar = 50 µm. The values are average of ±standard deviation.
OCSCs, ovarian cancer stem cells; PTX, paclitaxel; Sal, salinomycin.
*p<0.05.

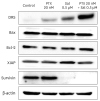
4. Effects of salinomycin on apoptotic signaling proteins in OCSCs
We examined whether salinomycin induced apoptosis by measuring the expression of DR5, Bax, Bcl-2, XIAP, and Survivin in OCSCs. The treatment combining salinomycin with PTX significantly increased DR5 and decreased Survivin. There was no change in the levels of Bcl-2, Bax, and XIAP (Fig. 4). These results suggest that salinomycin induced apoptosis through a non-mitochondrial pathway, consequently inhibiting cell growth.
Fig. 4
Effect of PTX and salinomycin on apoptosis-related proteins of OCSCs. Cells were exposed to PTX (20 nM) and salinomycin (0.5 µM) alone or in combination for 48 hours. After treatment, total protein was isolated. Expression of DR5, Bax, Bcl-2, XIAP, and Survivin proteins was analyzed by western blotting. β-actin was used as an internal loading control.
Bax, Bcl2-associated X protein; Bcl-2, B cell lymphoma 2; DR5, death receptor 5; OCSCs, ovarian cancer stem cells; PTX, paclitaxel; Sal, salinomycin; XIAP, X-linked inhibitor of apoptosis protein.
5. Induction of apoptotic cell death by salinomycin in combination with PTX
An apoptosis assay was performed to evaluate the cause of the inhibition of OCSC growth using annexin V-FITC/PI staining kit. Flow cytometric analysis showed that the combination of salinomycin and PTX dramatically increased the apoptotic cells upon 48 hours of exposure (Fig. 5A). Under the same conditions, activity of caspase-3, an executioner caspase of apoptosis, showed a 2.25-fold increase, and histone-associated DNA fragmentation showed a 4.67-fold increase (Fig. 5B, C).
Fig. 5
Effect of PTX and salinomycin in apoptosis. (A) After treatment with DMSO (control) or PTX (20 nM) and salinomycin (0.5 µM) alone or in combination, cultured OCSCs were harvested, fixed, stained with Annexin V/PI and analyzed by flow cytometry. (B) Cell lysates were prepared and used to profile caspase-3 activity. (C) DNA fragmentation was analyzed using an ELISA. The values are average of ±standard deviation.
Annexin V/PI, annexin V/propidium iodide; DMSO, dimethyl sulfoxide; ELISA, enzyme-linked immunosorbent assay; OCSCs, ovarian cancer stem cells; PTX, paclitaxel; Sal, salinomycin.
*p<0.05.

DISCUSSION
Despite the large number of individuals affected by gynecological malignancy, the mechanism involved in the pathology of ovarian cancer remains elusive. CSC theory offers a new perspective on tumor progression and cancer relapse. CSCs are characterized by self-renewal capability, differentiation potential, chemoresistance, and the ability to promote tumor proliferation. Previous research has found that a combination of salinomycin and metformin significantly inhibited the spheroid-forming capability of non-small cell lung CSCs, reflecting the suppression of self-renewal abilities [19].
Recent researchers have identified CD44 and CD117 as potential markers for the identification of cancer stem-like cells in ovarian malignancy [20]. Tumor spheroids generated from the ascites of ovarian malignancy patients are shown to have highly enriched CD44 and CD117 expressions. The CD44+CD117+ spheroid cells were resistant to chemotherapy and were able to initiate and serially propagate tumors in mice [21]. From analysis of 147 cases of epithelial ovarian cancers, Alvero et al. [22] found that the expression of CD44 was higher in metastatic tumors and tumor ascites. Subsequently, Luo et al. [23] reported that CD117+ lineage cells were capable of serial transplantation, asymmetric division, and the presence of these cells was correlated with chemoresistance. Another research group found that the combining salinomycin with doxorubicin increased the susceptibility of cancer cells and led to an intracellular accumulation of drugs [24]. In the current study, we present evidence of an antiproliferative and apoptotic effect of salinomycin in OCSCs. CSCs are known to have a greater spheroid-forming ability than cancer cells [6]. Our spheroid-forming assay demonstrated that the OCSCs treated with the combination of salinomycin and PTX presented the least amount of spheroid formation among the treatment groups, indicating that salinomycin has an antiproliferation effect on OCSCs.
OCSCs overexpress the stem cell–marking genes SOX2, OCT3/4, and NANOG, which are associated with high risk of ovarian cancer [2526]. Analyses from various studies regarding CSCs show that the stemness transcription factors play critical roles in stem cell proliferation and self-renewal [272829]. Since the gene expression of SOX2, and OCT3/4 represent strong CSC characteristics, a decrease in these gene expressions, as shown in the protein assay of treated OCSCs, indicates the antiproliferative effect of salinomycin on OCSCs.
SOX2 expression mediates resistance to carboplatin, cisplatin, and PTX, which indicates that it is a molecular driver of chemotherapy resistance in human ovarian carcinoma cells. CSCs inherently show high expression of SOX2, which indicates their apoptosis- and chemo-resistance [30]. In this study, salinomycin treatment lessened the expression of SOX2 and OCT3/4 mRNA and protein levels. Consequently, the down-regulation of SOX2 led to the reduction of the level of OCT3/4. We measured the effect of PTX on OCSCs after using the specific siRNA to silence SOX2, in order to determine whether reducing stemness can overcome the chemoresistance characteristic. We found that reducing SOX2 gave a nearly identical effect as treatment with combination therapy. It is not yet clear how salinomycin acts on OCSCs, but based on our finding, SOX2 could be a main protein affecting properties shared by CSCs. The underlying mechanism requires further investigation.
Previous research has shown that salinomycin induces extrinsic apoptosis in cisplatin-resistant OCCs [11]. In the present study, the results of several analytical methods, including caspase-3 activity, ELISAs of fragmented DNA, and annexin V-FITC/PI binding assays, demonstrated that salinomycin also induced apoptosis in OCSCs. This appeared to involve coordinated induction of elements through the death receptor pathway rather than the mitochondrial pathway. Protein expression of OCSCs treated with salinomycin and PTX showed up-regulation of DR5 and down-regulation of Survivin compared to the control sample, which indicates the apoptotic effect of salinomycin. Accordingly, salinomycin greatly enhances the efficacy of chemotherapeutics commonly used in the treatment of OCSCs.
Our results show that salinomycin in combination with PTX silences SOX2 expression and increases apoptosis of OCSCs. This evidence suggests the potential for salinomycin to be a component of effective ovarian cancer treatment.
 XML Download
XML Download