

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Epithelial ovarian cancer (EOC) is the most deadly of gynecological malignancies and the fifth most frequent cause of cancer death in women in developed countries [1]. High-grade serous ovarian carcinoma (HGSOC) is the most common and aggressive EOC histotype and accounts for the majority (60% to 80%) of EOC deaths [2]. Although in the last decades treatment has advanced significantly thanks to improved surgical techniques and chemotherapy regimens, the 5-year survival rate has not experienced substantial changes (between 35% to 40%) [13]. This poor overall prognosis is mostly a result of a combination of factors including the absence of distinctive symptoms and specific tumour markers on early stages so that most women are diagnosed at an advanced stage. Also, although the standard platinum-paclitaxel-based combination chemotherapy is initially successful, longer-term treatment is frequently compromised by the development of drug-resistant disease and toxicity profile [45]. Therefore, identification of novel prognostic and predictive markers in HGSOC that may help in treatment decision-making and eventually lead to the development of more effective therapies is of critical importance.
Over the last years, therapeutic strategies that inhibit DNA repair pathways or take advantage of intrinsic alterations in DNA-repair genes in tumors have been extensively considered as approaches to improve or predict efficacy of cancer therapies [6]. In particular, many different studies throughout the last two decades have evaluated in EOC and other tumour types the relation between cisplatin sensitivity and the components of the nucleotide excision repair (NER) system, which recognizes the DNA damage induced by platinum-based therapy [789]. Interestingly, Ceccaldi et al. [11], by using The Cancer Genome Atlas (TCGA) ovarian dataset, have recently described a subgroup of HGSOCs with NER alterations associated with a phenotype of platinum sensitivity similar to that of BRCA1/2 mutated tumors, with improved overall survival (OS) and progression-free survival (PFS) [11].
We previously showed that loss at 6q24.2-26 region is significantly associated with longer survival in HGSOCs independently of known prognostic factors [12]. By integrating copy number, RNAseq and survival data of 296 HGSOCs from TCGA, we demonstrated that the 6q24.2-26 deletion has an impact on gene expression, and among the genes that were significantly downregulated when lost, we defined several candidates, whose lower expression showed association with longer survival. One of the candidates proposed to explain the association of the 6q24.2-26 loss with survival is the General Transcription factor IIH Polypeptide 5 (GTF2H5, also known as TTDA). GTF2H5 encodes for an 8kD protein that forms part of the THFII complex, a multi subunit element involved in the local opening of the DNA double helix around DNA bulky lesions or interstrand adducts during the NER process [13141516]. The fact that GTF2H5 participates in DNA repair, especially in interstrand adducts removal [13] makes it a good candidate to perform additional studies in order to confirm its prognostic value and to examine its possible role in platinum-based chemotherapy sensitivity.
In the present study, we further explored the association found between survival of EOC patients and GTF2H5 both, at the protein expression level by using immunohistochemistry (IHC) in a series of 139 HGSOCs, and at the transcription level using publicly available independent datasets comprising altogether 855 HGSOCs. We also tested whether by including GTF2H5 we reinforced previous findings showing that NER alterations in HGSOCs are associated with improved outcome. Finally, in order to evaluate not only its prognostic, but also its potential predictive value, we have also performed functional experiments to assess whether stable short hairpin RNA (shRNA)-mediated GTF2H5 downregulation modulates cisplatin sensitivity in EOC cell lines.
MATERIALS AND METHODS
1. Patients and tumors
A total of 139 formalin fixed paraffin embedded primary HGSOCs were obtained from different hospitals throughout Spain after approval of the appropriate Human Research Ethics Committees at each participating center. Tumors were distributed in 5 tissue microarrays (TMAs) and 117 out of the 139 provided suitable GTF2H5 immunohistological staining comprising the final series evaluated in the present study (Supplementary Table 1). Eighty tumors were sporadic (with not reported first or second degree relative with breast or ovarian cancer) and 37 tumors classified as familial according to the criteria specified somewhere else [17]. All cases were blindly reviewed by three pathologists (IMR, JP and DH) and classified histopathologically. Histotype verification was assessed by H&E staining and evaluation of immunohistochemical markers such as Wilms tumour protein, tumour protein p53, estrogen receptor, progesterone receptor, and cyclin-dependent kinase inhibitor 2A. Grading was performed according to two-tier MD Anderson Cancer Center system [1819]. Most cases (72.4%) were of high International Federation of Gynecology and Obstetrics (FIGO) stage (stages III–IV using the FIGO system). All patients were treated according to a standardized protocol with a combination of taxane and platinum agents after debulking surgery. None of them had received neoadjuvant treatment. The length of OS was defined from the date of primary surgery to the date of patient death. PFS was calculated from the date of primary surgery to the date of disease progression, defined as an increase in cancer antigen 125 or radiological or surgical evidence of relapse. More detailed clinicopathological information is shown in Table 1. All the samples were obtained through The National Cancer Research Centre (CNIO) Biobank, which facilitates access to human samples for research purposes. CNIO Biobank was approved by the Spanish Law 14/2007 on Biomedical Research, and Royal Decree RD 1716/2011, and is registered in the National Registry of Biobanks with reference B.000848. The study was approved by Instituto de Salud Carlos III Research Ethic Committee CEI PI 24_2013.
Table 1
Clinicopathological features of patients and tumors

2. Immunohistochemical analysis
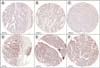
Morphology of tumors was reviewed on whole histological sections by gynecological specialized pathologists (JP, IMR, and DH) and cores were obtained from selected areas to build 5 TMAs as previously specified [20]. TMAs were designed to have two replicate cores for each tumour. Four-μm sections from TMAs were prepared with a semiautomatic microtome HM3508 (MICROM) and the tissue sections were deparaffinised and rehydrated in water. Antigen retrieval was performed in a DAKO PT Link in citrate buffer (pH, 6). Peroxidase activity was blocked with Dako Protein block for 10 minutes (containing casein 0.25% in phosphate-buffered saline) then incubated for 30 minutes with primary rabbit polyclonal anti-human anti-GTF2H5 antibody (1:100, 14539-1-AP, Proteintech, Chicago, IL, USA) for 30 minutes and detected with Dako Envision Plus kit (Dako, Glostrup, Denmark). Slides were counterstained with haematoxylin and coverslipped with DPX mountant for microscopy (VWR Int., Radnor, PA, USA). In agreement with the nuclear localization assigned to GTF2H5 in UniProtKB/Swiss-Prot according to experiments shown in Giglia-Mari et al. [16] immunohistochemical staining pattern of GTF2H5 was predominantly nuclear (Fig. 1). Scoring was exclusively based on the percentage of GTF2H5 stained nuclei within tumoural areas of each tumour regardless of staining intensity. In tumors with negative staining for GTF2H5, positively stained lymphocytes were used as internal positive control.
Fig. 1
Immunohistochemical analysis of GTF2H5 expression in high-grade serous ovarian carcinomas. Representative immunostaining of tumors belonging to the low- (A-C) and high- (D-F) GTF2H5 staining categories. Median stained nuclei in the series (5%) was used as threshold to define low (≤5%) and high (>5%) categories. Images correspond to tumour cores included in tissue microarrays.
3. GTF2H5 immunohistochemical staining and survival
The defined tumour categories of high- and low-GTF2H5 immunostaining were evaluated for their association with 5-year survival (OS). Five-year follow-up was chosen as survival endpoint in order to minimize the effect of non-ovarian cancer related deaths. Estimation of survival time distribution was performed using Kaplan-Meier (KM) method and statistical differences between curves were assessed with the log rank test. Univariate cox regression was used to estimate hazard ratios (HRs) and 95% CIs. The Cox regression model was also used for multivariate analysis. Time was censored at the date of last follow-up or at 5 years if alive, whichever came first. Statistical analyses were performed using SPSS ver. 17.0 (SPSS Inc., Chicago, IL, USA). All statistical tests were two-sided and nominal p-values less than 0.05 were considered statistically significant.
4. Validation at the global gene expression level
The association of GTF2H5 with survival was validated at the gene expression level using KM-plotter [21] with the JetSet probset. The KM-plotter is an online tool that allows the assessment of the prognostic value of the expression levels of microarray-quantified genes in ovarian cancer patients. We used the 2015 version of the database that includes gene expression data and survival information of 855 HGSOCs downloaded from Gene Expression Omnibus and TCGA (13 different datasets). Data was dichotomized at the automatically selected best fitted cut-off into GTF2H5-high and low expressing groups. KM and log-rank test were used to characterize the distribution and estimate the outcomes. In addition Cox proportional hazard model was used to estimate the HRs and 95% CIs. The association with 5-year OS and PFS was assessed in the 855 HGSOCs (grade 3) and in a more homogeneous group of 388 high FIGO (FIGO III and IV) with optimal debulking surgery HGSOCs.
5. Evaluation of NER alterations in TCGA dataset
Ceccaldi et al. [11] recently described a subset of tumors from TCGA ovarian cancer dataset with NER alterations (8% of the series) that were associated with improved OS and PFS [11]. Since the authors did not evaluate GTF2H5 we aimed to determine whether by reproducing the previously published analysis but with inclusion of GTF2H5 such associations were maintained or improved. As previously described [11] we accessed data for 316 patients with HGSOC in TCGA dataset for which promoter methylation, messenger RNA expression, DNA copy number and DNA sequencing information of exons was available [1122] and evaluated NER genes (XPA, XPC, DDB1, ERCC4, ERCC5, ERCC2, ERCC3, ERCC1, ERCC6, PCNA, ERCC8, LIG1, RAD23B, MNAT1, MMS19, RFC1, XAB2) for nonsynonymous or splice site mutations, promoter hypermethylation and homologous deletions concurrent with low mRNA expression levels. Since GTF2H5 does not shows mutations or bialellic deletions in TCGA dataset we evaluated those tumors that exhibit GTF2H5 low expression levels using stringent criteria (mRNA expression Z-score of less than –3). We then assessed associations with OS and PFS by using the KM method and log-rank test. Tumors that harboured BRCA mutation were not included in the survival analysis. As in Ceccaldi et al. [11], four samples with both, BRCA and NER mutations were also excluded.
6. Cell lines and cultures
Human ovarian cancer SKOV3 cells [23] were purchased from the American Type Culture Collection and cultured in RPMI 1640 medium with 10% fetal bovine serum (FBS), 1% penicillin/streptomycin, and 0.5% fungizone. COV504 cell line [24] was obtained from European Collection of Cell Culture and grown in Dulbecco's Modified Eagle's Medium with 10% FBS, 1% penicillin/streptomycin, and 0.5% fungizone. Cells were cultured at 37°C in a humidified 5% CO2 atmosphere.
7. shRNA mediated GTF2H5 silencing in ovarian cancer cell lines
shRNAs for human GTF2H5 (NM_207118) were obtained from Sigma-Aldrich (MISSION shRNA Bacterial Glycerol Stock, St. Louis, Mo, USA). Three different shRNA constructs were transduced and the one providing the best knockdown efficiency and non-cytotoxic effects was selected (SH3, TRCN0000062802) to stably-silence GTF2H5 expression in the SKOV3 and COV504 ovarian cell lines. A MISSION non-target shRNA vector (Scramble SHC002) was used as negative control. Briefly, lentiviral vectors were cotransfected with lentiviral packaging plasmids into 293FT cells (Invitrogen, Carlsbad, CA, USA) in the presence of Fugene 6 (Promega, Madison, WI, USA). The virus-containing supernatants were collected after 36 hours of transfection and filtered. Cells were infected and the stably transduced cells were selected by adding medium containing 1 mg/mL of puromycin. Silencing efficiency was tested by real-time quantitative polymerase chain reaction (qPCR) due to lack of available GTF2H5 antibody for Western blot analysis.
8. Real-time quantitative polymerase chain reaction
Total RNA was extracted from three independent plates for each cell line (RNAeasy Kit, Quiagen, Valencia, CA, USA) and converted to cDNA by using 500 ng as template (High Capacity cDNA RT kit, Applied Biosystems, Foster City, CA, USA). qPCR assays were designed for target gene (GTF2H5) and endogenous control (β-actin) using the Roche Universal Probe Library Assay Design Centre web site (Roche Applied Science, Indianapolis, IN, USA). Reactions were performed in triplicate using the ABI Prism 7900HT Sequence Detection System according to the manufacturer’s protocol (Applied Biosystems). Relative expression was determined using the qBase software that allows for PCR efficiency correction and implements normalization by endogenous gene.
9. WST-1 cytotoxicity assay
To evaluate the inhibition of cell viability by cisplatin treatment, 5×103 cells/well were pre-seeded 24 hours before treatment into a 96-well plate. Cells were treated with increased concentrations of cisplatin (0 to 80 μM) for 72 hours, and the WST-1 reagent was added to each well according to the manufacturer’s instructions (Roche Applied Science, Indianapolis, IN, USA). After 4 hours, cell viability was measured at an absorbance of 450 nm using a plate reader (Wallac 1420 VICTOR, PerkinElmer, Waltham, MA, USA).
RESULTS
1. IHC analysis of GTF2H5 expression
In order to further validate our prior finding of GTF2H5 association with survival [12], we analyzed GTF2H5 expression at the protein level by using immunohistochemistry in 139 HGSOCs assembled on 5 TMAs. A total of 117 tumour cores were interpretable for GTF2H5 staining, of which 107 (91.4%) had associated survival information. Uninterpretable staining was due to lack of enough tumour cells in the core or to complete core loss. Scoring of GTF2H5 was based on the percentage of positive tumoural nuclei regardless intensity. Values ranged between complete absence of staining and 100% stained cells with a median staining value of 5% in the series. High- and low-GTF2H5 staining categories of tumors were established according to the median staining value (high, >5%; low ≤5%). Fig. 1 shows representative examples of HGSOCs belonging to both categories. We evaluated whether GTF2H5 staining was associated with residual tumor size and other known prognosis markers such as FIGO stage, age of diagnosis, and immunohistochemical expression of the progesterone receptor [25] and we did not find statistically significant differences (p>0.05; Pearson chi-square test).
2. Low GTF2H5 IHC staining is associated with better survival in HGSOC
Association of GTF2H5 immunohistochemical expression with survival was assessed by KM analysis and log-rank test. We found significant differences in the 5-year survival between patients belonging to the GTF2H5-Low expression category (≤5% staining) and patients in the GTF2H5-High expression group (>5% staining; HR, 0.52; 95% CI, 0.29 to 0.93; p=0.024) with survival rates of 59% and 33.8%, respectively (Fig. 2A). Since ovarian cancer patients with BRCA1 or BRCA2 germline mutations have been shown to present better survival [26] and our series included familial ovarian cancer cases, we considered this variable as a possible confounder. We adjusted for this factor in the group of patients with known BRCA status (mutated or non-mutated) and the association remained statistically significant (HR, 0.19; 95% CI, 0.04 to 0.89; p=0.035).
Fig. 2
GTF2H5 expression is associated with longer survival in high-grade serous ovarian carcinomas (HGSOCs). (A) Kaplan-Meier survival curve showing a significant association between GTF2H5 immunohistochemical staining and 5-year survival. Patients with tumors belonging to the low staining group (≤5%) presented improved 5-year survival compared to the high-GTF2H5 staining group (>5%; p=0.024). (B-D) Kaplan-Meier curves from the Kaplan-Meier Plotter online tool. Lower transcriptional expression of GTF2H5 predicted enhanced 5-year survival in HGSOCs (p=0.023) (B) and in a homogenous group of high International Federation of Gynecology and Obstetrics (FIGO) stage optimally debulked HGSOCs (p=0.0007) (C). In high-FIGO stage optimally debulked HGSOCs there was also an association with improved progression-free survival (p=0.026) (D). The number of patients at risk at specific time points is shown below each diagram. The p<0.05 were considered to be statistically significant.

3. Decreased GTF2H5 expression levels are associated with better prognosis in a multi-institution HGSOC validation dataset
After finding significant differences in survival outcomes in our TMA set of samples stratified according to GTF2H5 IHC staining, we decided to extend the analysis to a larger and independent cohort of patients. To this end, we used the KM plotter, a free online tool that provides microarray gene expression data of 22, 277 genes in 855 HGSOCs from 13 different datasets [21]. In agreement with our IHC data we found that low expression of the GTF2H5 gene was associated with improved 5-year OS in 855 HGSOCs (HR, 0.80; 95% CI, 0.65 to 0.97; p=0.023) (Fig. 2B). To account for the effect of confounding factors other than histology and grade, we analyzed a more homogeneous group of 388 HGSOC of high FIGO stage (stage III and IV) and optimal debulking status, and the association was confirmed not only for improved 5-year OS (HR, 0.55; 95% CI, 0.38 to 0.78; p=0.0007) (Fig. 2C), but also for increased PFS (HR, 0.72; 95% CI, 0.54 to 0.96; p=0.026) (Fig. 2D). Altogether, these results suggest a significant prognostic relevance and a potential predictive value of GTF2H5 expression in HGSOCs.
4. GTF2H5 low expression reinforces the recently described association of NER alterations with improved PFS in HGSOCs
Upon the recent finding that HGSOCs from TCGA series with NER alterations are associated with improved OS and PFS [11], we decided to test whether by evaluating low GTF2H5-expressing tumors we maintained or improved such associations. With the criteria used in the previous published analysis, we found 24 of 316 tumors (8%) with NER alterations and also retrieved 9 additional tumors (3%) with low GTF2H5 expression using a stringent threshold (mRNA expression Z-score of less than –3) (Supplementary Fig. 1A). All NER alterations among themselves, but also in relation to GTF2H5 low expression, were mutually exclusive. Patients with tumors with NER alterations including those consisting of low GTF2H5 expression maintained a statistically significant higher median OS compared with patients with tumors without NER alterations and BRCA1/2 mutation (log-rank p=0.027). More importantly, in this comparison, the previously found trend toward statistically significantly higher median PFS in patients with NER alterations, reached significance by including low GTF2H5-expressing tumors (log-rank p=0.021) (Supplementary Fig. 1B).
5. GTF2H5 downregulation increases platinum sensitivity in HGSOC cell lines
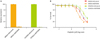
In order to further evaluate the potential predictive value of GTF2H5 expression levels, we aimed to determine the relationship between GTF2H5 deficiency and cisplatin sensitivity. For this purpose, by using specific shRNAs, we knocked down GTF2H5 in the ovarian cancer cell line SKOV3 and assessed cisplatin sensitivity. Knockdown efficiency of three different GTF2H5 shRNAs was tested by reverse transcription-qPCR (data not shown) and the construct that provided the highest efficiency was selected to carry out cytotoxicity assays (Fig. 3A). We observed a significant reduction in the half maximal inhibitory concentration (IC50) upon cisplatin treatment in the GTF2H5-silenced SKOV3 cells compared with control cells (transduced with scramble vector, p<0.05) (Fig. 3B). In order to further assess the generality of this result we also tested GTF2H5 silencing effect in COV504 cells (Fig. 3A). In this additional ovarian cancer cell model, a similar increase in cisplatin sensitivity was observed (Fig. 3B).
Fig. 3
GTF2H5 downregulation modulates cisplatin sensitization of ovarian cancer cells. (A) GTF2H5 expression in SKOV3 and COV504 ovarian cancer cell lines transduced with control (scramble) or GTF2H5-specific short hairpin RNA (shRNA). Data represent actin-normalized and relative to control mRNA expression levels of GTF2H5, as determined by real-time quantitative polymerase chain reaction. Data correspond to the mean of three independent experiments. (B) SKOV3 and COV504 ovarian cells transduced with control or GTF2H5-specific shRNA were treated with different concentrations of cisplatin for 72 hours and a standard WST-1 cytotoxicity assay was conducted to determine cell survival. A significant reduction in the half maximal inhibitory concentration (IC50) upon cisplatin treatment was observed in the GTF2H5-silenced SKOV3 cells compared with control cells (p<0.05). Consistent results were observed in GTF2H5-silenced COV504 ovarian cells. Values represent the SE of three independent experiments.
DISCUSSION
Less than 30% of ovarian cancer patients with advanced stage can be cured with the current standard treatment [27], which makes the definition of novel prognostic and predictive factors a priority in the field. Such markers may improve patients’ outcome by helping to select more appropriate therapeutic regimens and by providing insight into tumour biology that may eventually lead to the development of more effective targeted therapies.
Previous work carried out by our group demonstrated that a genomic loss of 6q24.2-26 region predicts better prognosis in HGSOC patients independently of known factors [12]. Identification of the deletion presents itself potential clinical value, as it could be used to guide the selection of patients towards more conservative therapeutic strategies in order to reduce side-effects and improve quality of life. However, further evaluation of the role played by the genes located in the lost region is essential to understand the possible mechanisms that drive the association and to reveal potential predictive significance. The NER-related gene GTF2H5 was among the candidates proposed to explain the association. GTF2H5 is a subunit of the transcription factor II human (TFIIH) complex, which is involved in the opening of the DNA double helix during the NER process. Nowadays the specific role of GTF2H5 in the complex remains controversial. Different studies suggest that GTF2H5 function could be focused on stabilization and protection of TFIIH from degradation [16] with a function presumed to make NER more efficient [28]. Other proposed its essential role in NER pathway and in embryonic development [29].
In the present study we found that low GTF2H5 protein expression defined by immunohistochemical staining is associated with improved 5-years OS in HGSOC patients. In addition, the analysis of 13 previously published transcriptomic datasets available in the KM-plot web tool [21] showed that HGSOCs with low-GTF2H5 expression levels present better 5-year OS than those with high-GTF2H5 expression levels. Altogether, these observations would confirm the prognostic value of GTF2H5 and upon validation in prospective studies this marker could assist in the selection of patients whose favorable outcome would support the use of new treatment regimens that aim to improve tolerability without jeopardizing efficacy. The possibility to evaluate GTF2H5 levels by using IHC on paraffin blocks is particularly interesting because this is the standard material routinely used for clinical diagnostic.
Besides the association with 5-year OS, our results also revealed an association of low-GTF2H5 expression with higher median PFS in a homogeneous group of optimally debulked high-FIGO stage HGSOC patients. PFS is a meaningful outcome measure of platinum response and this result supports the hypothesis that GTF2H5 might also have a predictive value for platinum-based therapy. Consistently, by including low GTF2H5-expressing tumors, the recent finding that patients with tumors with NER alterations present improved PFS [11] was reinforced. Also, shRNA-mediated GTF2H5-knock-down experiments showed a modest but significant cisplatin sensitization in ovarian cancer cell lines. Altogether, these findings are in agreement with the role of GTF2H5 in facilitating a more efficient NER [28] and in line with previous observations that cisplatin is generally more active in NER-compromised ovarian tumors [930].
Although we have been able to verify the individual prognostic value of GTF2H5 we should not overlook other candidate genes in the 6q24.2-26 region, such as ARID1B that forms part of the switch/sucrose non-fermentable (SWI/SNF) complexes that are involved in double strand break repair [3132] or the SNF2 histone linker PHD RING helicase SHPRH that plays a crucial role in an error-free repair of stalled replication forks caused by DNA damage [33]. It is plausible that the modest cisplatin sensitization effect observed by GTF2H5 silencing is additive or synergistic to the contribution of other DNA-repair related genes lost in the 6q24.2-26 deletion. Interestingly, a recent high-throughput data-based study defined 6q15-27 among eight DNA copy number losses that significantly affect candidate pathways associated with chemoresponse in EOC [34].
In conclusion, we have shown that low protein and expression levels of the NER-related gene GTF2H5 are associated with enhanced prognosis in HGSOC. In addition we provide evidence that GTF2H5 downregulation may also contribute to cisplatin sensitization. Our results warrant prospective validation in order to be translated into clinical practice and support further evaluation of the specific and combined effect of other candidates at 6q24.2-26.
 XML Download
XML Download