

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Colorectal cancers are the second most common cancer in males and third most common cancer in females in Korea. The incidence of colorectal cancers in 2010 was about 12.8% of all cancers. The incidence rate of colorectal cancers has continued to increase in both sexes in Korea [1].
Main treatment modalities for colorectal cancers are surgery, radiotherapy, and chemotherapy. Chemotherapy is frequently used as the main regimen in most advanced colorectal cancers. Oxaliplatin, a third-generation platinum complex, is one of the most effective chemotherapeutic agents in advanced colorectal cancers. It differs from cisplatin (1st generation platinum complex) in lack of nephrotoxicity and from carboplatin (2nd generation platinum complex) in mild hematological toxicity. However, it causes serious dose-related side effects such as peripheral neuropathy. The acting mechanism causing oxaliplatin to show stronger cytotoxicity to cancer cell with resistance to the other platinum compound drugs and the reason it is more effective when combined with other anticancer drugs are not known [2]. However, repressing expression of survivin that suppresses apoptosis was reported to be one of the mechanisms of oxaliplatin [3,4].
But tumor resistance to chemotherapy (chemo-resistance) presents a major cause of failure in chemotherapy [5]. So, many researchers have been searching for effective chemosensitizing agents. Many potential chemosensitizers are natural agents such as resveratrol. Resveratrol, one of the main constituents of red wine, is a natural agent with potential as an effective chemo-sensitizers [6].
Resveratrol can sensitize tumor cells to many chemotherapeutic agents by overcoming one or more mechanisms of chemoresistance, including survivin down regulation [7,8]. Resveratrol inhibits the growth of a wide variety of tumor cells in culture [9]. Resveratrol has been shown to potentiate the cytotoxic effects of paclitaxel, cisplatin and gemcitabine [10-12]. But in some tumor cells, resveratrol has been shown to act as cytoprotector. Expression of survivin in cancer cells decreased even at low doses of resveratrol, but the effects were dose-dependent [8].
Some cell survival proteins were associated with development of chemoresistance, including c-FLIP [13], X-linked inhibitor of apoptosis [14], and survivin [15]. Survivin is highly expressed in human cancers, including colorectal cancers, but not detectable in nonproliferating normal adult tissues [16]. The role of survivin in the pathogenesis of cancer is inhibition of apoptosis, regulation of the mitotic spindle checkpoint, promotion of angiogenesis and chemoresistance [17]. And high survivin level is related to poor prognosis and resistance to therapy, radiation and chemotherapeutic agents [18]. Thus, survivin is a promising target for sensitization of cancer cells to radiation and chemotherapeutic agents [15].
Our previous studies show that oxaliplatin can suppress survivin expression of colon cancer cells. In order to maximize oxaliplatin therapy, we examined whether resveratrol, a natural phytochemical known to have chemopreventive effects on cancers, can have a chemosensitizing effect upon cotreatment with oxaliplatin. Contrary to our expectation, we found that survivin suppression by YM155 or oxaliplatin pretreatment can be nullified by resveratrol posttreatment. Resveratrol can have a recovering effect on oxaliplatin-mediated cytotoxicity in HCT116 cells.
METHODS
Cell line and reagents
HCT116 human colon cancer cells were obtained from Korean Cell Bank (Seoul, Korea). Cells were cultured in Roswell Park Memorial Institute (RPMI) 1640 medium supplemented with 10% fetal bovine serum (FBS), 100 U/mL penicillin and 100 µg/mL streptomycin in a humidified incubator at 37℃ in an atmosphere of 90% air and 5% CO2.
Oxaliplatin was obtained from Sanofi-Synderabo (Seoul, Korea), diluted in sterile phosphate buffered saline (PBS) and stored at 4℃. Penicillin, streptomycin, RPMI 1640, FBS, resveratrol, crystal violet, Hoechest 33258 and YM155 were purchased from Sigma (Sigma-Aldrich Co., St. Louis, MO, USA). MTT kit for cell proliferation was purchased from Promega (Fitchburg, WI, USA). The experimental concentration of oxaliplatin was used according to our previous study [4].
Antibodies against survivin (FL-142, sc-10811), Bcl-2 (100, sc-509), caspase-3 (H-277, sc-7148) and Erk2 (sc-154) were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA).
Growth inhibition assay
Inhibition of cell growth by oxaliplatin, resveratrol and YM155 was assessed by 3-(4,5-dimethylthiazol-2yl) 2,5-diphenyltetrazolium bromide (MTT) assay. After pretreatment with oxaliplatin (2, 5 µM) or resveratrol (30, 50 µM) for 48 hours, the surviving cells were harvested and posttreated with oxaliplatin and resveratrol separately or in combination for 48 hours.
Cells were pretreated with YM155 (1 mM) for 1 hour and medium was exchanged with new fresh media containing oxaliplatin, resveratrol, and a combination of oxaliplatin and resveratrol and maintained for 48 hours.
Healthy growing HCT116 cells were detached by trypsin-ethylenediaminetetraacetic acid (EDTA) treatment and 2.5 × 104 cells per ml were resuspended in RPMI containing 10% FBS, which were then seeded into 96-well culture plates with six duplicates. After 24 hours, the culture was maintained for an additional 48 hours in the absence (control) or presence of oxaliplatin, resveratrol, and YM155 as described in the figure legends. After 48 hours culture, 20 µL of 5 mg/mL stock of MTT was added into each well. After 1 hour incubation, culture media was removed and 0.1 mL of dimethyl sulfoxide was added into each well to dissolve the formazan crystals. The intensity of color was measured at 570 nm. All values were normalized to control wells.
Crystal violet staining
Fifty percent confluent HCT116 cells in 24-well plates were treated with oxaliplatin, resveratrol, and in combination of oxaliplatin and resveratrol for 24 hours. Cells were then fixed in 10% buffered formalin, washed with PBS, incubated with 1% crystal violet (Sigma-Aldrich Co.), solubilized with 1% Triton X-100 and analyzed for absorbance value at 590 nm on a Tecan GENios Plus (Tecan Group Ltd., Zurich, Switzerland).
Apoptosis and morphologic assay
HCT116 cells were cultured with oxaliplatin (2, 5 µM), resveratrol (30, 50 µM), and oxaliplatin with resveratrol. After 24 hours incubation, cells were stained with Hoechest 33258 and observed in fluorescent microscope (IX71, Olympus Co., Tokyo, Japan). Morphologically apoptotic cells were counted in four randomly selected different sections of each culture.
Reverse transcription-polymerase chain reaction
Total RNA was isolated from the cells (1×106 cells/sample) using Trizol reagent (Life Technologies, Rockville, MD, USA). The following primer sequences were used for polymerase chain reaction (PCR) amplifications of survivin: (forward: 5'-CCACCGCATCTCTACATTCA-3' and reverse: 5'-TATGTTCCTCTATGGGGTCG-3'), survivin-2B ( forward: 5'-CCACCGCATCTCTACATTCA-3' and reverse: 5'-AGTGCTGGTATTACAGGCGT-3'), survivin-ΔEx3 (for ward: 5'-CCACCGCATCTCTACATTCA-3' and reverse: 5'-TTTCCTTTGCATGGGGTC-3'), GAPDH (forward: 5'-TGGACCTGACCTACGGCAACAGATA-3' and reverse: 5'-GCCCTCATACTGGAAACCC-3') genes.
Cells that were cultured were dissolved in 1 mL of cold Trizol solution and lysed. Then, they went through the centrifuge at 12,000 g at 4℃ for 10 minutes, and the supernatant was moved into a new eppendorf tube and was then settled at room temperature for 5 minutes. Chloroform, 200 µL, was added, thoroughly agitated for 15 seconds, and centrifuged at 12,000 g at 4℃ for 15 minutes after being settled for 3 minutes at room temperature. Supernatant-containing RNA was carefully moved to a new eppendorf tube, and 500 µL of isopropyl alcohol was added and centrifuged at 12,000 g at 4℃ for 10 minutes after being settled at room temperature for 10 minutes. Supernatant was removed after checking the presence of RNA clusters, and 1 mL of 75% ethanol was added to clean it. Then, it was centrifuged at 7,500 g at 4℃ for 5 minutes, ethanol was removed, and the RNA was obtained after drying it in the air for 15 minutes. This RNA was dissolved in secondary distilled water that was treated with diethyl pyrocarbonate (DEPC, Sigma Chemical Co.), and the concentration of RNA was measured with a spectrophotometer measuring the optical density at 260/280 nm. The separated RNA was stored at -80℃.
In creating the cDNA, 2 µL of Oligo dT (Promega) and secondary distilled water that was treated with DEPC were added to 5 µg/µL of the RNA obtained from the HCT116 colon cancer cell strain until the mixture was 10 µL. Then, a fixation reaction was allowed to occur for 10 minutes at 70℃, the mixture was then cooled with ice for 10 minutes, and a master mix (5 × first strand buffer, 4 µL; 0.1 M dithiothreitol, 2 µL; 2.5 M deoxyribonucleotide triphosphates (dNTPs), 2 µL; RNase inhibitor, 1 µL) was added. The mixture was reacted for 2 minutes at 42℃, 1 µL of reverse transcriptase (superscript II [Life Technologies] was added, and the mixture was reacted again for 50 minutes at 42℃ and 15 minutes at 70℃ to finally obtain the cDNA). Secondary distilled water without nuclease, 36.5 µL, and 5 µL of master mix (10 × buffer [500 mM KCl, 100 mM Tris-HCl {pH 8.3}, 15 mM MgCl2]), 4 µL of dNTP, 1 µL of primer sense, 1 µL of primer antisense, and 0.5 µL of Taq DNA polymerase (TaKaRa Taq, Takara, Shiga, Japan) were added to 2 µL of cDNA. Then, the mixture was placed in a DNA thermal cycler (GeneAmp PCR system 9700, Applied Biosystems, Foster City, CA, UAS) for 2 minutes at 94℃, 1 minute (denaturing) at 94℃, 1 minute (annealing) at 60℃, and 1 minute (extension) at 72℃. Under the conditions of PCR, the process was repeated 28 times to amplify the reaction. After the PCR reaction, electrophoresis was executed in 1.5% agarose gel, and GAPDH was modified by using the same method.
Western blot analysis
The cells were lysed in lysis buffer (50 mM Tris, 100 mM NaCl, 2.5 mM EDTA, 1% Triton X-100, 1% Nonidet P-40, 25 µg/mL aprotinin, 25 µg/mL leupeptin, 25 µg/mL pepstatin A, and 1 mM phenylmethysulfnyl fluoride). After centrifugation at 10,000 g for 15 minutes, the supernatant was used for Western blot analysis. Protein concentration was determined by the Bio-Rad Protein Assay kit (Bio-Rad, Hercules, CA, USA). Aliquotes of cell lysates containing 50 µg of protein were separated by 3% sodium dodecyl sulfate-polyacrylamide gel electrophoresis. Following electrophoresis, proteins were transferred electronically onto polyvinylidene fluoride membranes (Bio-Rad). Membranes were incubated for 1 hour at room temperature with blocking buffer, TBS-T (20 mM Tris, pH 7.6, 100 mM NaCl, 0.1% Tween-20) and 5% nonfat dry milk with gentle agitation. After washing the membranes with TBS-T three times, membranes were incubated overnight at 4℃ in TBS-T buffer containing 5% nonfat dry milk and with one of the following antibodies (1:1,000 dilution): survivin (sc-10811), Bcl-2 (sc-509), caspase-3 (sc-7148). The membranes were washed three times with TBS-T and incubated with secondary antibodies (1:5,000 dilution) in TBS-T buffer containing 5% nonfat dry milk for 2 hours at room temperature with gentle agitation. The membranes were washed again three time with TBS-T, the protein bands were visualized by enhanced chemiluminescence detection system (Amersham, Little Chalfont, UK). The reacted bands were detected with an image analyzer (LAS-2000, Fuji Film, Tokyo, Japan).
Membranes were stripped in a stripping buffer containing 100 mM2-mercaptoethanol, 2% sodium dodecyl sulfate, 62.5 mM Tris-HCl, pH 6.7 at 55℃. The membranes were reprobed for the level of ERK-2. All Western blots were performed at least 3 times for each experiment.
RESULTS
Resveratrol posttreatment nullifies oxaliplatin pretreatment induced survivin suppression
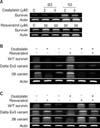
Survivin mRNA expression is suppressed with oxaliplatin treatment (5 µM) for 2 days. However, when we treated the cells with resveratrol (30, 50 µM for 2 days incubation), survivin mRNA expression was not suppressed (Fig. 1A). With surviving cells upon oxaliplatin pretreatment (5 µM, for 2 days), we posttreated with resveratrol (50 µM), oxaliplatin (5 µM), and combination of resveratrol and oxaliplatin for 2 days. Survivin mRNA expression was suppressed with preoxaliplatin treatment. However, when resveratrol was posttreated on the surviving cells, the survivin mRNA expression was restored. This restoration effect was also detected in survivin delta Ex3, 2B variants (Fig. 1B).
And a reciprocal experiment was performed in this setting. When we posttreated with oxaliplatin on the surviving cells with resveratrol pretreatment, survivin mRNA expression was suppressed. However, when we posttreated with resveratrol or oxaliplatin + resveratrol on these cells, the survivin mRNA expression was restored. We also observed restoration of survivin delta Ex3, 2B variants. When we pretreat HCT116 cells with resveratrol, oxaliplatin-mediated surviving suppression was restored (Fig. 1C).
Resveratrol posttreatment nullifies YM155 and oxaliplatin pretreatment induced cell proliferation suppression
We pretreated HCT116 cells with YM155, a specific survivin inhibitor to suppress the survivin expression for 1 hour. After YM155 pretreatment, cells were treated with oxaliplatin, resveratrol, and a combination of oxaliplatin and resveratrol for 24 hours. Cells were then stained with crystal violet and analyzed for viability of cells by measuring absorbance at 590 nm. Resveratrol restored cell viability upon YM155 mediated survivin suppression. This restoration effect of resveratrol also occurred in combined pretreatment of YM155 and oxaliplatin as shown in Fig. 2.
Resveratrol posttreatment nullifies YM155 and oxaliplatin pretreatment induced apoptotic activity
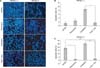
We also stained the treated cells with Hoechst to assess the apoptotic cells. Resveratrol showed a cytoprotective effect restoring from undergoing apoptosis. Oxaliplatin induced apoptosis. However, when cells were treated with oxaliplatin + resveratrol, apoptotic cells were markedly decreased (Fig. 3A, B). When we inhibited survivin expression by YM155, resveratrol treatment protected cells from undergoing apoptosis even in combined treatment with oxaliplatin (Fig. 3C). Resveratrol had a cytoprotective effect on HCT116 cells restoring from undergoing apoptosis induced by YM155 and oxaliplatin. Combined treatment with oxaliplatin and resveratrol abolished oxaliplatin-mediated apoptotic induction.
Resveratrol posttreatment nullifies YM155 and oxaliplatin pretreatment induced protein expression.
Oxaliplatin pretreatment suppressed expression of survivin, Bcl-2, caspase-3 protein levels in HCT116 cells. YM155 abolished expression of survivin, bcl-2, and calspae-3 protein levels additively upon oxaliplatin cotreatment. However, resveratrol posttreatment restored expression of survivin, Bcl-2, caspase-3 protein levels down-regulated by oxaliplatin. Decreased expression of bcl-2 and caspase-3 protein levels by YM155 and oxaliplatin seems to be related to survivin suppression but expression of bcl-2 and caspase-3 protein levels was restored with combined treatment with resveratrol (Fig. 4). Taken together, resveratrol may directly induce restoration of survivin suppressed by oxaliplatin and YM155 pretreatment. Importantly, resveratrol may play a role as a cytoprotector in survivin suppressed cells in some cancer cells.
DISCUSSION
Oxaliplatin, a third-generation platinum complex, is the one of the most effective chemotherapeutic agents for the treatment of advanced stages of colorectal cancers in clinic. The advantage of oxaliplatin therapy over other platinum compound drugs is stronger cytotoxic activity on platinum refractory cancers but the mechanism of action is not well established [2]. One of the potential mechanisms of oxaliplatin superiority could be due to down-regulation of survivin expression upon oxaliplatin treatment [3,4].
Our data have shown that oxaliplatin can suppress survivin expression in both transcripts and protein level of HCT116 cells. Alternative splicing variants of survivin (2B, deltaEx3) were also suppressed upon oxaliplatin challenge. Furthermore, pretreatment of YM155, a small molecule known to specifically inhibit survivin, additively suppressed survivin expression of HCT116 cells upon oxaliplatin challenge. YM155 pretreatment and oxaliplatin change additively suppressed protein expression level of Bcl-2 and caspase-3 of HCT116 cells. Taken together, combinatorial treatment of YM155 and oxaliplatin may increase cytotoxic effects on colon cancers. Thus, we propose that combination treatment of oxaliplatin and YM155 could be a promising treatment modality for advanced stages of colon cancers in clinics.
Chemotherapy is the main regimen in most advanced colorectal cancers to prevent recurrence or metastases. But tumor resistance to chemotherapy (chemo-resistance) presents a major cause of failure in chemotherapy [5]. Also, chemotherapeutic agents have considerable side effects. Because of chemoresistance and side effects, many researchers have been searching for effective chemosensitizing agents to potentiate the chemotherapeutic effect and reduce the side effects. Many phytochemicals have been studied for chemosensitization activity. Resveratrol, one of the main constituents of red wine, is a natural agent found to be an effective potential chemo-sensitizers [6].
Resveratrol (3, 4', 5-trihydroxy-trans-stilbene) was first isolated in 1940 and has anti-infective, antioxidant, and cardioprotective functions [19]. In 1997, resveratrol was found to have anticancer potential [20] and the anticancer activities are mediated through modulation of several cell signaling molecules that regulate cell cycle progression, inflammation, proliferation, apoptosis, invasion, metastases, and angiogenesis of tumor cells [9,21,22].
Most of the reports indicate that resveratrol sensitize tumor cells to chemotherapeutic agents by modulating cell survival proteins. Resveratrol potentiated apoptosis induced by chemotherapeutic agents by down-regulating survivin expression and increasing apoptosis [7]. Resveratrol has also been shown to sensitize tumor cells by inducing caspase activation. It was reported that resveratrol may affect many intracellular signaling molecules including suppression of survivin. But in some cancer cells resveratrol may act with a cytoprotection effect.
Due to the chemosensitizing effect of resveratrol, we initially anticipated that resveratrol cotreatment with oxaliplatin may additively suppress survivin expression of HCT116 cells. Upon resveratrol cotreatment with oxaliplatin, however, there was no additive suppression of survivin in HCT116 cells. More surprisingly, we observed that resveratrol challenge on oxaliplatin pretreated HCT116 cells can nullify oxaliplatin-mediated survivin suppression and restore survivin expression of HCT116 cells in both transcripts and protein level. Oxaliplatin challenge on resveratrol pretreated HCT 116 cells, however, cannot nullify oxaliplatin-mediated survivin suppression of HCT116 cells. Taken together, our results clearly showed that resveratrol has a strong recovering effect on oxaliplatin-mediated cellular toxicity. To further examine resveratrol-mediated recovering effects, we performed cell proliferation and apoptosis assays. Oxaliplatin pretreated HCT116 cells, which underwent suppression of cell proliferation activity and induction of apoptosis, were able to restore their cellular viability upon resveratrol challenge. Furthermore, YM155 pretreated HCT116 cells, which underwent survivin suppression in both transcripts and protein level, were able to restore their survivin expression levels upon resveratrol challenge. In Western blot analysis, we were able to show restoration of survivin, BCl-2, and caspase-3 protein levels of HCT116 cells upon resveratrol post-treatment on YM155 and oxaliplatin pretreated HCT116 cells.
But unlike its proapoptotic action in tumor cells, resveratrol has antiapoptotic effect on chondrocytes [23], and inhibit p53-dependent apoptosis [21].
Expression of survivin in cancer cells decreased even at low doses of resveratrol, but the effects were dose-dependent [8].
Effects of resveratrol were dependent on cell, tissue type and concentration.
Survivin constituted the fourth top "transcriptome" in colorectal, lung, brain, and breast cancer by genome-wide searches [24]. This reflects an "oncofetal" pattern of expression because survivin is highly expressed in embryonic organs, is nearly undetectable in most differentiated normal tissue, and becomes dramatically re-expressed during tumor development [25,26].
Survivin participates in the cellular stress response, in which binding of the protein to the molecular chaperone heat shock protein 90 helps tumor cells cope with unfavorable environments and other exogenous stresses and thus preserve cell proliferation and cell viability [27,28].
Inhinition of survivin by the Eli-Lilly survivin specific synthetic single stranded DNA-antisense oligonucleotides (LY2181308) resulted in significantly increased apoptosis, and decreased clonogenic survival when combined with radiation and also suggested that survivin shows a radioresistance factor and molecular biomarker, predicting tumor response in patients with rectal cancer treated with neoadjuvant radiochemotherapy [29].
The role of survivin in the pathogenesis of cancer is inhibition of apoptosis, regulation of the mitotic spindle checkpoint, promotion of angiogenesis and chemoresistance [17].
And high survivin level is related to poor prognosis and resistance to the therapy, radiation and chemotherapeutic agents [18]. So attention has been given to survivin as a target for sensitization of cancer cells to radiation and chemotherapeutic agents [15].
In this paper, resveratrol may act as cytoprotective agent in HCT116 colon cancer cells. Especially, resveratrol can restore survivin suppression induced by oxaliplatin or YM155, specific survivin inhibitor. Also, it protects the HCT116 colon cancer cells from apoptosis by oxaliplatin and YM155.
We recommend cautious use of resveratrol as a chemoprevention or chemosensitizing agent though resverstrol's chemosensitizing effect is in the process of being establishment by many other studies.


 XML Download
XML Download