

 ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Pancreatic cancers possess a poor prognosis with less than 4% of five-year survival rate. Along with a global increase in the number of pancreatic cancer patients, they account for 4th-5th cause of death induced by cancers. The incidence has increased more than 2 folds in Korea as compared to 10 years ago and pancreatic cancers are the 5th cause of death among the cancers in organs [1]. Although radical resection rate seems to be improved lately because of sufficient pancreatic resection, extensive lymph nodes and retroperitoneal plexus dissection, and combined resection of major blood vessels; long-term survival rate is not as high as expected [2]. In addition, several therapeutic methods have been carried out as the treatments of unresectable pancreatic cancers, such as anticancer drug perfusion therapies via the hepatic arteries and portal veins, chemoradiation therapies, and radiation therapies during surgery along with systematic chemotherapy and external irradiations.
The pathway of apoptosis is roughly classified into 2 types including the receptor-mediated (extrinsic) apoptosis through death receptors (death receptors, Fas/FasL, tumor necrosis factor-α, etc.) and the mitochondria-mediated apoptosis induced by various factors such as chemotherapeutic agents and reactive oxygen species. Additionally, 3rd pathway which is caused by endoplasmic reticulum (ER) stress has been suggested recently [3]. Caspase, a cystein protease which is activated in all kinds of cells during apoptosis, plays the most significant roles in apoptosis, even though such stress possesses distinct cell signaling pathways activating apoptosis. Casepase-8 protease is mainly activated in the receptor-mediated apoptosis pathway while caspase-9 protease is in the mitochondria-mediated apoptosis pathway. Meanwhile, caspase-12 protease that is known to be activated by the increase of intracellular calcium concentration is involved in the reticulum pathway [4]. Furthermore, the activated caspase-12 protease directly activates caspase-9 protease so that induces apoptosis eventually [5]. Endoplasmic reticulum is an important organelle for protein syntheses and folding that secretions, cell membranes, and golgi apparatus are organically associated with lysosome as well as for various other functions including glycosylation of new proteins, disulfide bonds, and protein folding and assembly. Additionally, ER is a storage of intracellular calcium ions either pumping calcium ions via sarcoplasmic/ER Ca2+-ATPase or releasing calcium ions by inositol 1,4,5-triphosphate thereby playing crucial roles in homeostatic maintenance.
Quercetin belongs to polyphenolic flavonoids abundant in fruits, vegetables, and various plants with various biological as well as physiological effects including antioxidant activity, anticancer activity, antiviral activity, apoptosis induction, protein kinase C inhibitory activity, cell cycle modulation, angiogenesis inhibition, and angiotensin converting enzyme inhibitory activity. Especially, quercetin inhibits the release of P-glycoprotein and increases the anticancer activity against adriamycin in in vitro breast cancer cell lines [6]. Multiple studies have investigated regarding the anticancer activity of quercetin; in particular, they have reported that quercetin at various dosage levels suppresses cancer cell growth in various cell lines such as breast cancers, colorectal cancers, stomach cancers, head and neck cancers, leukemia, lung cancers, melanoma, and ovarian cancers [6-8]. However, induction of apoptotic signaling or signaling molecules has not been clearly understood. Therefore, the objective of this study was to investigate apoptosis mechanisms and functions of the signaling molecules of ER stress when quercetin was administered in pancreatic cancer cell line in order to provide effective treatments of pancreatic cancers.
METHODS
Materials
Cell line
PANC-1 cells, a human pancreatic cancer cell line, was purchased from Korean cell line bank (Seoul National University) and subcultured throughout the experiment.
Reagents and equipment
Dulbecco's modified eagle medium (DMEM), antibiotics, trypsin, and fetal bovine serum (FBS) were purchased from Gibco BRL Co., (Grand Island, NY, USA) while culture dish (24-well plate and 10-cm dish) was obtained from Falcon Co., (Becton Dickinson, San Jose, CA, USA). Quercetin, 3-[4,5-dimethylthiazol-2-yl]-2,5-diphe-nyltetra zolium bromide (MTT), 4'-6-diamidino-2-phenylindole (DAPI), bicinchoninic acid, dimethyl sulfoxide (DMSO), reduced glutathione, and N-acetyl-L-cysteine were purchased from Sigma Co., (St. Louis, MO, USA).
Methods
PANC-1 cell line culture
PANC-1 cells were cultured in a CO2 incubator (37℃, 5% CO2) with DMEM containing 10% FBS and culture medium was replaced every 24 hours. Then, quercetin was administered in the cells at log phase followed by the biochemical experiments relevant to apoptosis. Quercetin was dissolved in 100 mg/mL of DMSO and then diluted in culture medium.
Measurement of cell survival rate
Cell survival rate was measured using MTT analysis. Cells (1 × 105 cells/mL), 1 mL, were aliquot on a cell culture plate (24-well plate) and stabilized in a CO2 incubator for more than 12 hours. Then, samples were treated based on the conditions accordingly. MTT solution (5 mg/mL in PBS), 1/10 of culture solution, was added to the culture cells. The culture solution was eliminated 4 hours after the treatment and 1 mL of DMSO was added to dissolve the cells. Then, absorbance was measured at 540 nm utilizing spectrophotometer (Thermo max, Columbia, MD, USA). Cell survival rate was expressed as percentage (%) by comparing with the control group.
DAPI staining
The cells treated with quercetin were fixed in 4% formaldehyde solution and washed with PBS. After staining with 10 µM of DAPI for 10 minutes, they were washed with PBS again and then the changes of nucleic morphologies were observed via a fluorescence microscopy (MPS60, Leica Microsystems Welzlar GmbH, Bensheim, Germany).
Western blot analysis
Cultured PANC-1 cells were collected and then washed two times with cold PBS. Obtained cells were reacted with the homogenization solution (HEPES, 50 mM; pH, 7.4; NaCl, 150 mM; deoxycholate, 1%; ethylenediaminetetraacetic acid [EDTA], 1 mM; phenylmethylsulfonyl fluoride, 1 mM; aprotinin, 1 µg/mL) at 4℃ for 30 minutes. Equal amount of the homogenization solution (200 µg) and 2 X sample buffer were mixed and then subjected to a heat treatment at 100℃ for 3 minutes to denature proteins followed by 12.5% and 15% sodium dodecyl sulfate-polyacrylamide gel electrophoresis. After the gel electrophoresis, the proteins on the gel were transferred to a nitrocellulose membrane using an electrotransfer system (0.8 mA/cm) (Ellard Inc., Seattle, WA, USA) and then incubated in the blocking buffer (5% skim milk) at room temperature for 2 hours. Primary antibodies against Bcl-XL, Bak, Grp78/BiP, phospho-PERK, PERK, ATF6, phospho-eIF2α, eIF2α, GADD153/CHOP, and β-actin were diluted to 1:1,000 in Tris-buffered saline (TBS-T) containing 0.05% (v/v) of tween-20 and then incubated with the nitrocellulose membrane at room temperature for 2 hours. Antirabbit IgG conjugated horse radish peroxidase (HRP) and anti-mouse IgG conjugated HRP, the secondary antibodies against the primary antibodies, were diluted in TBS-T (1:3,000) and incubated at room temperature for 1 hour. Then, they were developed utilizing enhanced chemilluminescence kit (Amersham, Buckinghamshire, UK).
Measurement of intracellular active oxygen
Formation of intracellular active oxygen was determined using 2',7'-dichlorofluorescein diacetate (DCF-DA, Sigma Co.), a fluorescent probe. DCF-DA, a nonfluorescent substance, converts to fluorescent green DCF in the presence of peroxides associated with intracellular hydrogen peroxides. Quercetin was administered to PANC-1 cells and then cultured in 5 µM DCF-DA at 37℃ for 30 minutes prior to the cell harvest. Cultured cells were washed with PBS (pH 7.4). They were treated with 1% trypsin-EDTA and then harvested. The cells were washed with PBS again and fluorescence was measured utilizing flow cytometry (FACSCalibur, BD Biosciences, San Jose, CA, USA). The analysis using CellQuest software (BD, Franklin Lakes, NJ, USA) was followed afterward.
Cell cycle analysis
Cell DNA was strained with propidium iodide (PI) and then the cell cycles were measured using flow cytometry (FACSCalibur). After treating the cells with quercetin, they were collected and washed twice with PBS. DNA of the washed cells (1 × 106) was subjected to incubation in 600 µL of PI solution (Triton X-100, 0.1%; PI, 20 µg/mL; RNase, 200 µg/mL) for 20 minutes. Based on the information obtained from sub-G0/G1 of the flow cytometry, the cell cycles were analyzed utilizing CellQuest software (BD).
Measurement of intracellular calcium accumulation
Fluo 3-AM (Molecular Probes, Eugene, OR, USA), a fluorescent probe, was applied to determine intracellular calcium accumulation caused by quercetin. After treating quercetin to PANC-1 cells, the cells were cultured with 5 µM of Fluo-3 AM in a CO2 incubator for 30 minutes before the cell harvest. Harvested cells were washed with PBS (pH 7.4) followed by fluorescence measurement via flow cytometry (FACSCalibur). Intracellular calcium accumulation was analyzed utilizing CellQuest software (BD).
RESULTS
The effects of various anticancer drugs on cell survival rate of PANC-1 cells
To investigate the effects of various anticancer drugs utilized in cancer treatments on the cell survival rate of PANC-1 cells, adriamycin, cisplatin, gemcitabine, and 5-fluorouracil (5-FU) were treated at various dosage levels and then the cell survival rate was determined by MTT assay. When adriamycin and cisplatin were treated, dose-dependent apoptosis of PANC-1 cells were observed at the high dosage levels of more than 5 µg/mL and 2 µM (Fig. 1A, B) while apoptosis of gemcitabine, which was widely utilized in the treatments of pancreatic cancers generally, was not exhibited even at 125-2,000 µg/mL (Fig. 1C). When 5-FU was treated, the cell survival rate started decreasing at 6.25 µg/mL and then 76% of the cell survival rate was observed at 50 µg/mL (Fig. 1D).
The effects of quercetin on survival rate of various cancer cells
To identify the effects of anticancer drugs of quercetin, cytotoxicity of variously cultured cancer cell lines were confirmed using MTT assay. Various dosage levels of quercetin were treated first and then cultured for 24 hours. In results, a decrease of dose-dependent cell survival rate was exhibited in HL-60, HepG2, Hep3B, and MCF-7 cells whereas the cell survival rate of H9c2 cells was not changed even at 50 µg/mL of quercetin (Fig. 2).
Changes of PANC-1 cells survival rate by quercetin
To evaluate the cytotoxicity of PANC-1 cells by quercetin, quercetin was treated at various dosage levels for 24 hours and then the changes of the cell survival rate were measured by MTT method. The PANC-1 cells treated at 12.5 µg/mL, 25 µg/mL, 50 µg/mL, and 100 µg/mL of quercetin showed 76%, 62%, 43%, and 18% of cell survival rate of the control group, indicating dose-dependent reductions (Fig. 3A). When 50 µg/mL of quercetin was treated for various time periods, time-dependent cytotoxicity was found with reduced survival rate from 12 hours after the treatments (Fig. 3B).
Histologic changes of PANC-1 cells by quercetin
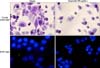
To confirm that the apoptosis by quercetin was mediated by apoptosis mechanisms, crystal violet staining and nucleic DAPI staining which allowed observing morphologic characteristics of apoptosis were carried out. PANC-1 cells that were treated with 50 µg/mL of quercetin for 24 hours were stained and then observed via a phase contrast microscopy and a fluorescence microscopy. As results, the quercetin treated group showed a notable decrease in the number of cells along with overall cell condensation as compared to the control group. The nucleus of the control group showed an oval shape with intact and homogenous fluorescence in the DAPI staining while fluorescent fragments that were segmented into several pieces were observed in the quercetin treated group. At the same time, reductions in the number of cells as well as morphologic changes were accompanied (Fig. 4)
Changes of capspase protease activity in PANC-1 cells by quercetin
To investigate whether PANC-1 apoptosis by quercetin is related to caspase protease activity, the enzymatic activity of caspase-3 and caspase-9 proteases were measured using fluorescent substrates. Caspase protease activity was determined in the homogenized solution of PANC-1 cells treated with 50 µg/mL of quercetin by time. In results, the activity of caspase-9 protease started to increase from 3 hours after the quercetin treatment and then was increased more than 5.8 folds at 6 hours compared to the control group (Fig. 5A). In addition to the caspase-9 protease activity, the activity of caspase-3 protease started elevating from 18 hours after the quercetin treatment and then showed more than 7.2 folds increased activity at 24 hours compared to that of the control group (Fig. 5B). Such results implied that quercetin elevated the activity of caspase-9 protease in PANC-1 cells and induced caspase-3 protease activation in downstream of signal transduction.
Effects of apoptosis of PANC-1 cells by quercetin on the changes in mitochondrial membrane potential
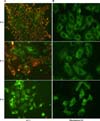
To identify the changes in mitochondrial membrane potential, JC-1 and rhodamine 123 fluorescent staining was carried out. After treating PANC-1 cells with 50 µg/mL of quercetin for various time periods, they were stained with JC-1 and then observed via a fluorescence microscopy. As results, the quercetin treated PANC-1 cells, an orange color as like the control group, turned to a fluorescent green (Fig. 6A).
In case of rhodamine 123 staining, the mitochondria of PANC-1 cells without the quercetin treatment showed a fluorescent green with a punctuated-pattern and were localized in a part of cytoplasm. In contrast, the mitochondria with the quercetin treatment converted into a diffused-pattern and were evenly distributed throughout the entire cytoplasm. Such results suggested that the quercetin treatment caused the changes in mitochondrial membrane potential of PANC-1 (Fig. 6B).
Effects of quercetin on the changes of Bcl-XL and Bak protein expression in PANC-1 apoptosis
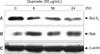
To evaluate the changes of Bcl-XL and Bak protein expression when treating with quercetin, PANC-1 cells were treated with 50 µg/mL of quercetin and then western blot was carried out. In results, the expression of Bcl-XL protein, possessing antiapoptotic functions, was decreased from 6 hours after the quercetin treatment (Fig. 7A) while the expression of proapoptotic Bak protein was increased with the same treatment (Fig. 7B). Through β-actin, it was confirmed that the protein content was the same (Fig. 7C).
Changes in intracellular calcium of PANC-1 cells by quercetin
To confirm the calcium ions that were accumulated within the cells in the apoptosis of PANC-1 cells by quercetin, the cells were stained with Fluo-3 AM dye and observed via flow cytometry. As results, it was observed that the calcium ions were accumulated in the cells from 6 hours after 50 µg/mL of quercetin treatment and showed a time-dependent calcium increase with the maximum accumulation at 24 hours (Fig. 8).
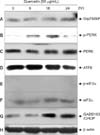
Effects of quercetin on the changes in the proteins associated with ER stress pathways of PANC-1 cells
Since we confirmed that the accumulation of calcium ions within the cells was increased, the changes in the protein expression relevant to ER stress pathways were investigated in order to elucidate the pathways. The cells were treated with 50 µg/mL of quercetin for 6, 18, and 24 hours. The proteins of the collected cells were separated and then subjected to western blot. An increased of Grp78/BiP protein was observed 24 hours after the quercetin treatment (Fig. 9A). Phosphorylation of PERK protein started increasing from 6 hours after the quercetin treatment and achieved to the maximum activity at 12 hours. Then, it tended to decrease gradually but no changes were exhibited in the PERK protein expression (Fig. 9B, C). Additionally, ATF6 protein, the down-stream signaling protein of PERK, was decreased from 18 hours after the quercetin treatment (Fig. 9D) and phospho-eIF2α and eIF2α didn't show any changes in their expression (Fig. 9E, F). In case of GADD153/CHOP, the expression was increased from 18 hours after the quercetin treatment and then tended to keep increasing until 24 hours (Fig. 9G). Through β-actin, it was confirmed that the protein content was the same (Fig. 9H). The results implied that quercetin went through the ER stress pathways which induced the increase of GADD153/CHOP protein expression finally by increasing Grp78/BiP protein as well as activating PERK protein in PANC-1 cells.
Antitumor exacerbation effects of quercetin in PANC-1 cells
Antitumor exacerbation effects of PANC-1 cells were investigated when the combined treatments between various anticancer drugs utilized in the treatment of pancreatic cancers and quercetin were administered. Combinations of either gemcitabine or 5-FU with quercetin were treated and then stained with PI dye after 24 hours. The degree of apoptosis was observed via flow cytometry. The subG0/G1 fraction in the control group, 7%, was increased by 12% with 50 µg/mL of 5-FU treatment only, 13% with gemcitabine treatment only, 14% with the treatment of 5-FU and gemcitabine combined, and 16% with quercetin treatment only, respectively.
However, when the combinations of either 5-FU or gemcitabine with quercetin were treated, subG0/G1 fraction was increased by 15% and 19%, respectively. Especially, when quercetin, gemcitabine, and 5-FU were treated at the same time, 25% of the subG0/G1 faction was found, indicating a noted increase of apoptosis (Fig. 10).
DISCUSSION
In recent years, diagnosis and treatments of cancers have improved greatly. However, pancreatic cancers that the incidence has been increasing globally showed poor prognosis as there are difficulties in early diagnosis and treatments so that many studies still left to be investigated in this area. There are several factors inducing pancreatic cancers such chemicals, virus infection, high calorie diets, excessive intake of saturated fatty acids, smoking, alcohol intake, chronic pancreatitis, diabetes, and genetic factors [6,7]. Pancreatic cancers are one of the main diseases with the 5th highest death rate by cancer. Pancreatic cancers possess the highest fatality after diagnosis among all cancers. Approximately 70-80% of the pancreatic cancer patients who are diagnosed in the early stage without symptoms experiences recurrences of the cancers after surgery. Therefore, not only local treatments like surgery but also chemotherapy is important in the treatments of pancreatic cancers. In pancreatic cancers treatments, anticancer therapies inhibit cancer development and relieve the symptoms in the patients thereby improving the quality of the patients' life. They further extend their lifespan ultimately [1]. It is known that 5-FU is an effective and basic drug for pancreatic cancer treatments. It has been widely utilized either alone or combined with other drugs such as gemcitabine in general. Since gemcitabine has been introduced to the pancreatic cancer treatments, 5-FU is specially used as secondary treatments in the patients who didn't experience the effects of gemcitabine, instead of being utilized as primary treatments. Gemcitabine (20, 20-difluorodeoxicytidine), a pyrimidine antimetabolite, is similar to deoxycytidine. There are several drug toxicities involved in gemcitabine such as bone marrow suppression, lethargy, cold symptoms, nausea, vomiting, and edema. Utilization of gemcitabine only showed superior chemotherapy effects especially in advanced pancreatic cancers compared to that of 5-FU pancreatic cancers so that it has been established as a standard of anticancer therapies. Moreover, it is reported that gemcitabine is effective even in the cases where 5-FU didn't show potency for pancreatic cancers in the past [9]. However, its effects including extension of lifespan have not achieved a fully satisfied level so that many efforts have been made to enhance anticancer effects by combining gemcitabine with other anticancer drugs. In the present study, it was observed that gemcitabine and 5-FU didn't have significant cytotoxicity in PANC-1 cells. In particular, gemcitabine at a high dosage (2 mg/mL) didn't show the differences in cell survival rate whereas 5-FU induced the decrease of cell survival rate as similar to high dosage adriamycin or cisplatin.
Apoptosis, an important mechanism to maintain homeostasis of cells or organs, has developed in the area of cell biology since late 1980s. The importance of cell formation, differentiation, and functions has been known especially in nervous systems and immune systems [10]. Abnormalities of apoptosis mechanisms are critical in the understanding of cancer occurrence, resistance expression against cancer treatments, the occurrence of autoimmune diseases, and the pathologic phenomena by degenerative diseases and human immunodeficiency virus [11]. Many anticancer drugs including ara-c, cis-platinum, cyclophosphamide, adriamycin, etoposide, teniposide, vincristine, mitoxantrone, taxol, hydoxyurea, and bleomycin induce apoptosis in various cancer cell lines [12]. Fas/Fas L system, sphingomyelin/ceramide pathway, the activation of early immediate genes, caspase family cysteine protease, and endonuclease are involved in the action mechanisms [13]. However, pancreatic cancer cells possess low reactivity against anticancer drugs and are resistant to apoptosis toxicity by anticancer drugs compared to other cells [14]. Apoptosis, the planned cell death, is one of the essential physiological phenomena for normal organ development as well as homeostasis maintenance of tissues in multicellular organisms; and many biological phenomena such as rapid cell dehydration due to cell contractions, blebbing of cell membranes, the increase of calcium concentration in the cytoplasm, chromosome condensation, segmentation of the DNA 'ladder' shape due to endonuclease activation, transglutaminase activation, nucleus breakages, and the formation of apoptotic body are accompanied [15]. Whereas surgical treatments are well known as pancreatic cancer treatments, the mechanisms of anticancer drug resistance have not been clearly elucidated yet. Therefore, the authors investigated the effects of quercetin on apoptosis induction in PANC-1 cells and the signal transduction mechanisms. As results, quercetin was sensitive to PANC-1 cells induced a decrease of cell survival rate in a time- and dose-dependent manner. We confirmed that the action mechanism was the apoptosis characterized by chromatin condensation and nucleic acid fragmentation with reductions in the number of cells and morphological changes. In particular, the toxicity against H9c2 myocardial cells which are widely known as cardiac dysfunction mechanisms, one of the side effects occurring in chemotherapy treatments, was not exhibited (Fig. 1).
Quercetin is included in flavonoids present in plants that are used as foods. It possesses useful pharmacological effects due to its strong antioxidant effects such as prevention of circulatory diseases, anti-inflammatory effects, and anticancer effects [16]. The anticancer effects of quercetin are associated with the mechanism that suppresses mutations occurred by substances like t-butyl hydroperoxide by eliminating free radicals [17]. In addition, quercetin showed excellent pharmacological effects in circulatory systems including suppression of myocardial cell damages and arteriosclerosis prevention [18]. Moreover, it relaxes vascular smooth muscles resulting in lowering blood pressure or inhibiting arrhythmia [19]. It is also effective in other diseases that oxidative stresses are involved such as suppression of the damages by ultraviolet ray [20] and antidiabetic effects [21]. In this study, quercetin showed a significant cytotoxicity against liver cancer cell lines like HepG2 and Hep3B as well as breast cancer cell line like MCF-7; but not against H9c2 which was originated from myocardial cells. Such results are in agreement with the study performed by Park et al. [22], addressing that quercetin protects the myocardial cell damages due to oxidative damages.
Additionally, quercetin activated caspase-9 protease and caspase-3 protease in PANC-1 cells and there were differences in mitochondrial membrane potential observed via JC-1 and rhodamine 123 fluorescent dyes. Among Bcl-2 family proteins involved in the loss of mitochondrial membrane potential in apoptosis, we confirmed the reduction of Bcl-XL and increased expression of Bak. However, further studies may need to clarify whether the results of this study are in agreement with previous studies [23] regarding either intracytoplasmic release of cytochrome c relevant to caspase-9 protease or the signal transduction mechanisms via the activation of downstream caspase protease. Calcium ions play important roles in intracellular signal transduction [24]. In nerve cells and excitatory cells, the calcium ion tunnels at cell membranes are opened when it is depolarized and then calcium ions are flew in through the tunnels thereby increasing the concentration of intracellular calcium ions. Also, the stored calcium ions in the cells are released by free inositol 1,4,5-triphosphate by the stimulation from several receptors so that the concentration of intracellular calcium ions are elevated as well [25]. In nonexcitatory cells, the concentration of intracellular calcium ions is also increased in the presence of stimulation due to hormone etc. For many years, multiple studies in various fields have investigated the functions of such changes in the concentration of calcium ions [26]. Intracellular calcium ions in the cells are generally maintained at very low concentration because the calcium ions are actively stored in ER which is one of the storages of intracellular calcium [27]. High calcium ion concentration in ER is essential for the functions of ER as like new protein folding and disulfide bond formation. If calcium ion homeostasis is not maintained properly in ER, the functions of ER are lowered. In the present study, the increase of intracellular calcium ion concentration due to quercetin was confirmed by flow cytometry through Fluo-3 AM staining so that the ER stress pathway seemed to be activated.
The process of ER stress response is activated to recover and maintain the functions of ER; and it is classified into 4 steps. In the first step, activated PERK reduces the transitions of protein synthesis through phosphorylation of eIF-2 α (α subunit of eukaryotic initiation factor 2) so that new protein synthesis is decreased [28]. Then, the second step induces ER chaperones such as BiP in order to enhance protein folding in ER. The third step called ER stress associated degradation is to degrade the unfolded or mis-folded proteins in ER through intracytoplasmic ubiquitin-proteasome system. Lastly, when the functions of ER are not able to be recovered due to the severe ER stress-induced stress, apoptosis pathways are activated resulting in the elimination of damaged cells. In ER stress condition, the 4 response steps are implemented in a time-dependent manner [29]. When quercetin was treated in this study, the expression of Grp78/BiP protein, phosphorylation of PERK protein, and the expression of GADD153/CHOP protein were increased in PANC-1. Rodrigues et al. [30], reported that the expression of Bcl-2 family protein was controlled by CHOP protein due to ER stress pathways. The results of the present study that quercetin induced the increase of Bak protein are in agreement with the results from Rodrigues et al. [30], addressing that calcium ions were released in ER. In this study, however, the expression of ATF6 was reduced and eIF 2 α activity as well as expression didn't change. Therefore, it is considered that PANC-1 apoptosis by quercetin is the ER stress pathways via PERK-GADD153/CHOP signal transduction pathways; but further studies are requested to identify signal transduction mechanisms accurately.
Taken all together, quercetin represented a superior dose- and time-dependent cytotoxicity in PANC-1 cells compared to that of gemcitabine and 5-FU, the chemotherapeutic agents for pancreatic cancer treatments, and induced nucleus fragmentation and apoptosis by caspase-3 protease activity. In addition, quercetin induced a decrease of Bcl-XL expression and an increase of Bak expression resulting in functional changes of mitochondria. This may contribute to caspase-9 protease activity. Quercetin caused the accumulation of intracellular calcium ions in PANC-1 cells. It also induced ER stress and further apoptosis mechanisms based on an increase of Grp78/BiP expression, activation of PERK protein, and an increase of GADD153/CHOP expression. Quercetin accelerated the apoptosis effects especially when combined with gemcitabine or 5-FU. Thus results in the study suggest that quercetin could be utilized as a cancer treatment by increasing the sensitivity against pancreatic cancer cells with anticancer drugs resistance.
The results of this article could be summerized follow as:
Quercetin induced dose- and time-dependent cell death in PANC-1 cells.
Quercetin induced fragmentation of nucleic acids in PANC-1 cells.
Quercetin activated caspase-9 protase and caspase-3 protase in PANC-1 cells.
Quercetin lowered the expression of Bcl-XL and increased the expression of Bak in PANC-1 cells thereby decreasing the mitochondrial membrane potential.
Quercetin induced the accumulation of intracellular calcium ions and the activation of the ER stress pathways based on the increase of Grp78/BiP expression, PERK activation, and the increase of GADD153/CHOP expression.
When quercetin was combined with gemcitabine or 5-FU, the cell death was accelerated in PANC-1 cells.
In conclusion, quercetin activated apoptosis mechanisms via intracellular caspase protase activity, mitochondria dysfunction, and ER stress in PANC-1 cells, the human pancreatic cancer cell line. It also increased the reactivity of pancreatic cancer cells against anticancer drugs so that facilitated apoptosis.


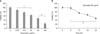



 XML Download
XML Download