

 ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Tacrolimus (FK506) is an immunosuppressive drug, discovered in 1984 from Streptomyces tsukubaensis, that is used to prevent rejection of transplanted organs following the organ transplantation [1-4]. FK506 suppresses T cell activation by inhibiting calcineurin and the calcineurin-dependent transcription factors, the nuclear factor of activated T cells (NFATc), which are the central regulators of the T cell function. However, the molecular mechanisms underlying how FK506 regulates NFATc in T cell activation are obscure. Previous studies have indicated that FK506 efficiently blocks proliferation of T cells by inducing a G0/G1 cell cycle arrest.
MicroRNAs (miRNAs) are small RNAs that regulate the expression of complementary messenger RNAs. Hundredsof microRNA genes have been found in diverse animals, and many of these are phylogenetically conserved [5-7]. Recently, microRNAs have been shown to target particular sets of mRNAs, thereby preventing translation or accelerating mRNA turnover [5,6,8,9]. Therefore miRNAs play important roles in normal regulation of gene expression for developmental timing, cell proliferation, and apoptosis.
This study investigated the Jurkat T cell line expresses cytotoxicity when treated with different concentrations of FK506, and analyzed the expression pattern of miRNA when stimulated by FK506 using the miRNAs microarray, as well as the expression pattern of a gene that is related to the differentiation, activation and proliferation of T cells after being affected by the change of miRNAs. Furthermore, by analyzing the relationship between the expression pattern of the genes related to apoptosis and the biological pathway, this study was expected to provide the new approach to the biological effect of FK506.
METHODS
Jurkat human T-lymphocyte lines obtained from the Korean type Culture collection (Seoul National University, Seoul, Korea) were maintained in RPMI-1640 (Gibco BRL, Grand Island, NY, USA) tissue culture medium supplemented with 10% fetal calf serum at 37℃ with 5% CO2. FK506 was solubilized at a concentration of 5 mg/mL in dimethylsufoxide, maintained at 20℃, and diluted in the RPMI-1640 to the final concentrations of 0.1 to 100 mol/L. All cell cultures were incubated at 37℃ with 5% CO2. Cell viability was measured using a 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazoliumbromid assay at 24 and 48 hours after FK506 treatment.
RNA extraction
RNA was extracted from cell lines using a standard Trizol (Invitrogen, Carlsbad, CA, USA) protocol, and modified by washing the final RNA pellet with 70% EtOH. The cell lines were sectioned on a cryostat prior to homogenization in Trizol. RNA quantity and quality were assessed by Nanodrop (Genomic Tree, Daejeon, Korea) and Agilent 2100 bioanalyzer (Agilent Technologies, Santa Clara, CA, USA), respectively.
MicroRNA labeling and hybridization were done by using Cy3-dyed total RNA. Our miRNA microarray (Agilent's Human genome 4 × 1.5 K glass slide) contained a probe for 534 mature human microRNAs and 64 mature viral microRNAs (Sanger miRBase release 9.1).
Unbound sample was removed from the beads by washing with 1× TE and resuspending in 1× tetramethylammonium chloride buffer after the hybridization. Streptavidin-phycoerythrin, premium grade (Invitrogen) was added to the beads (1:100 dilution) and incubated for 10 minutes at 50℃ to bind to biotin moieties on the cDNA. Samples were scanned by the laser confocal scanner (G2565BA, Agilent Technologies) and median fluorescence intensity values were acquired using the Feature extraction software (Agilent Technologies).
- Amplification and labeling: performed using the Agilent's Low RNA Input Linear Amplification kit PLUS.
- Microarray hybridization: performed using the Agilent's Gene Expression Hybridization Kit.
- Microarray wash: used the Agilent's Gene Expression Wash Buffer Kit.
- Scan and image analysis: used the Agilent's DNA micro array scanner and the Feature Extraction Software.
The experiments were repeated three times each. The analysis of the microarray result used the Agilent's GeneSpring Software to sort the miRNAs that showed increase or decrease by normalization, clustering and filtering. And to identify the function of the target genes of these miRNAs, DAVID's program from the website was used to analyze the biological function.
RESULTS
As a result of treating Jurkat cell line with various durations and concentrations of FK506, 10 µM of FK506 lowered the viability of the cell to 92.2% after 24 hours of treatment, 77.3% after 48 hours, 62.5% after 72 hours, and 46.2% after 96 hours. With 20 µM of FK506, the viabilities were 87.2% after 24 hours, 68.4% after 48 hours, 45.7% after 72 hours, and 22.45% after 96 hours. Also, 30 µM of FK506 showed 78.08%, 56.52%, 33.49%, and 12.91% viability at above mentioned durations, respectively. Therefore, the treatment with FK506 decreased the viability of human Jurkat T-cell in dose- and time-dependent manners. Thus in this study, the cell line was treated with 20 µM of FK506 for 72 hours, the combination of which shows roughly 50% viability, for the microRNAs microarray analysis.
Change in the microRNA expression by the effect of FK506 during apoptosis of Jurkat cell
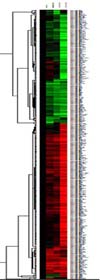
MicroRNA array was performed to the experimental group of Jurkat cell lines, which were each treated with 20 µM FK506 for 24, 48, and 72 hours, and the changed signal intensity was analyzed after eliminating the background signal. The groups of miRNAs that were expressed more than twice, or less than a half compared to the control group were classified and marked as a cluster tree (Fig. 1). The group of miRNAs that were expressed more than twice the control group was marked red, and the group that showed the expression of less than a half the control group was marked green, and thereby sorted the miRNAs according to the pattern of expression in a time-dependent manner.
Analysis of the miRNAs that decreased in expression during FK506-mediated apoptosis of Jurkat cell
As a result of the microarray performed to the Jurkat cells treated with 20 µM FK506 for 24, 48, and 72 hours, the number of miRNAs that decreased in expression to less than a half compared to the control group was 21 in total, and by searching for any relationship between these miRNAs and the genes, the miRNAs miR-19a, -19a*, -19b, -20a*, -106a, -142-5p, -301a, -301b, and -487a were found at the end of mRNA of NFATC3 (NFAT4) gene, which regulates the apoptosis of Jurkat cell mediated by FK506 (Table 1, Fig. 2). Also, the entire genes that are regulated by these miRNAs were analyzed using DAVIDs bioinformatics, and it was found that the genes regulated by these microRNAs, which showed decreased expression, were related to the biological processes, such as p53 signaling pathway and the regulation of cell cycle (Table 2).
Analysis of the microRNAs that increased in expression during FK506-mediated apoptosis of Jurkat cell
As a result of the microarray performed to the Jurkat cells treated with 20 µM FK506 for 24, 48 and 72 hours, the number of microRNAs that showed increased expression to more than twice of the control group was 18 in total, and by searching for any relationship between these microRNAs and the genes, the microRNAs miR-486-5p and miR-518c* were detected at the end of mRNA of TP53I3 gene, which is related to triggering p53 associated cell death. As for FKBP8 gene, the intracellular calcineurin inhibitor, the microRNAs miR-330-5p and miR-631 were detected (Table 3, Fig. 3).
Also by using DAVIDs bioinformatics, the group of genes that are regulated by these 18 miRNAs were found to be associated with various biological processes, including calcium signaling pathway (Table 4).
DISCUSSION
Most of the studies regarding FK506 have focused primarily on its function as an inhibitor for the intracellular calcium-dependent calcinuerin and the drug's target protein, NFAT, which is a transcription factor of T cells, as well as its function to regulate the expression of cytokines. But not enough studies were conducted regarding the role of FK506 to express the genes related to apoptosisand its mechanism.
According to the recent studies, the intervention of miRNAs in RNA molecules plays an important role in regulating the gene expression, and is phylogenetically conserved. The gene expression is regulated by disassembling or inhibiting the mRNA of the target gene with very small RNAs [6]. Hundreds of miRNAs are investigated in nematodes, plants and animals through computer analysis and Reverse Transcriptase-Polymerase Chain Reaction cloning [10,11]. Hence this study used Jurkat human T lymphocyte line to verify that the cell's viability decreases in a time and dose dependent manner when treated with FK506, the immunosuppressive drug that is used after the organ transplantation to prevent rejection.
In order to analyze the change cause by FK506 in the expression pattern of miRNAs and the signal pathway stimulated by the drug's target molecule, 20 µM of FK506, which shows about 50% of cell viability, was treated for 24, 48, and 72 hours and followed by miRNA microarray using the total RNA to analyze the expression of miRNAs associated with the activation of T cell by FK506 and the drop in cell viability.
As a result of treating 20 µM of FK506, 21 kinds of miRNA including miR-20a* decreased to less than a half in timedependent fashion, and other 18 kinds of miRNAs including miR-518c* increased to more than twice of the control group. Most of the decreased miRNAs are related to the immunosuppressive functions of FK506, including inhibition of the well-known cytokines, such as interleukin 2 and CD40 [12,13], and the regulation of calcium/calmodulin dependent kinases [14-16] and NFATs [17], which are the intracellular transcription factors. FK506 is known to release FKBP12, which is bound to the transforming growth factor (TGF)-β receptor in a chronic lymphocytic leukemia B cell, and phosphorylates and activates the receptor, thereby causing phosphorylation of Smad and leading to apoptosis [18,19]. In a smooth muscle cell, however, it was found that the activation of the TGF-β receptor by FK506 stimulates cell growth [1,18]. Ataxia telangiectasia-mutated (ATM) kinase is a multifunctional enzyme that is activated after birth and known to take part in suppressing the onset of ataxia-telangiectasia (AT) [20,21]. The cells that lack ATM show markedly increased activation of thiol redox and DNA synthesis compared to a normal cell. The regulation of ATM signal transduction by FK506 affects the cell cycle by controlling the intracellular thiol redox pathway and therefore is involved in suppressing the abnormal DNA synthesis and oncogenesis [22,23]. On the other hand, the target genes of the miRNAs that are increased by FK506 were identified and their biological functions were analyzed using DAVIDs bioinformatics. The analysis detected numerous genes that are associated with the intracellular energy metabolism pathway, the production of nucleotides; pyrimidine and purine, the production of amino acids related to protein synthesis, the calcium signal transduction pathway and the signal transduction by T cell receptors. Benedini et al. [24], reported that in case of lung transplant patients, the immunosuppressive drugs increase metabolism of glucose and leucine. It was also reported that when FK506 was treated to a mouse after the liver resection for regeneration, the insulin receptors in the liver cell of mouse increased in number, and the energy metabolism and the amino acid synthesis were boosted [25,26]. It was also recently reported that calcineurin controls the synaptic plasticity and the transcription factor, NFAT in a synapse of a neuron. Furthermore, it was found that the dendritic branching could increase in number by suppressing the calcineurin molecule [27,28]. The FK506-binding proteins (FKBP12 and 12.6) are involved in a regulation of ryanodine receptors that control the calcium ion in the sarcoplasmic reticulum of a heart, and the treatment of FK506 loosens their binding and calcium is released from the sarcoplasmic reticulum as a result [29]. In addition, when IP3R is inhibited in association with the control of neurite outgrowth by calcium, the neurite outgrowth was suppressed in the early stage, whereas the inhibition of calcium-induced calcium release by ryanodine receptor suppresses the neurite outgrowth in the later stage. And when treated with FK506, the neurite outgrowth was suppressed in the early stage as well [28].
In conclusion, as a result of treating FK506 to a Jurkat cell line and running the miRNA microarray, it was found that FK506 not only took part in the suppression of T cell proliferation/activation by inhibiting calcineurin in Jurkat apoptosis, but also affected the miRNAs that are involved in the regulation of various signal transduction pathways that are related to the causes of apoptosis and the cardiovascular diseases, kidney problems, neurotoxicity, diabetes, and the acute hearing impairment triggered by a long-term use of FK506.






 XML Download
XML Download