

 ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Immunological suppression is a well recognized consequence of trauma and hemorrhagic shock, and contributes to infectious complications, ultimately leading to sepsis and multiple organ dysfunction syndrome (MODS) [1]. There are several mechanisms of post-traumatic immune impairment, including T cell dysfunction. T cell dysfunction after traumatic stress is characterized by a decrease in T cell proliferation. The addition of prostaglandin E2 (PGE2), which depresses immune function after hemorrhage and trauma, to T cells decreases T-cell proliferation, while hypertonic saline (HTS) restores PGE2-induced T-cell suppression [2].
Recently, it has become apparent that macrophage migration inhibitory factor (MIF) plays a central role in several immune responses, including the modulation of several cytokines and T-cell proliferation [3,4]. By controlling immune and inflammatory responses, MIF is thought to play an important role in the pathophysiology of septic shock and chronic inflammatory diseases [5-7]. The role of MIF mediating HTS restoration of T cell dysfunction is unknown.
Therefore, we hypothesize, that T cell immune restoration by HTS occurs, at least in part, by a MIF-mediated mechanism. The proposed experiments will provide the first evidence supporting MIF as the primary mechanism for restoring T cell dysfunction by HTS.
METHODS
Cells culture and cell stimulation
Jurkat cells clone E6-1 (ATCC, Manassas, VA, USA) were maintained in Roswell Park Memorial Institute-1640 (Invitrogen, Carlsbad, CA, USA) supplemented with 10% fetal bovine serum, 2 mM glutamine, 10 mM 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES), 100 U/mL penicillin/streptomycin at 37℃ in a 5% carbon dioxide incubator. Cells were cultured to a density of 5 × 105 cell/mL. Cell viability, as determined by tyropan blue dye exclusion, was >99%. For the protein extracts, cells were plated at a density of 2.5 × 106 cell/mL in a 6-well flat-bottom culture plates, and were stimulated with PGE2 (1 µg/mL) (Sigma-Aldrich Co., St. Louis, MO, USA) in the presence or absence of HTS at 20 mmol/L (HTS20) and 40 mmol/L (HTS40) above isotonicity, resulting in sodium concentrations of 160 mmol/L and 180 mmol/L, which were measured by a GEM Premier 3000 (Instrumentation Laboratory, Lexington, MA, USA), respectively.
Protein extracts
After incubation for 24 hours at 37℃, the cells were washed 2 times in cold phosphate buffered saline (PBS) and then centrifuged for 10 minutes. Cells pellet were resuspended in 10 µL per 2 × 106 cell/mL of superlysis buffer (protease inhibitors, 1 M HEPES, 5 M NaCl, 0.5 M ethylenediaminetetraacetic acid, 1 mM NaOV4, 20% Triton X-100, 50 mM phenylmethylsulfonylfluoride), incubated on ice for 7 minutes, and then centrifuged at 3,000 × g for 15 minutes at 4℃. The supernatant was then transferred to an eppendorf tube and used for assay. The total protein concentration was determined by the Bradford method using a commercially available assay kit (Thermo Fisher Scientific, Rockford, IL, USA) [8]. The prepared protein lysates were aliquoted and used for Western blot analysis.
T-cell proliferation assay
Jurkat cells were plated in 96-well flat-bottom tissue culture plates to attain a final concentration of 2.5 × 106 cell/mL. 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-tetrazolium bromide (MTT) were conducted by the quantity of PGE2 0.01, 0.1, 1, 10 µg/mL and MTT assay value was the lowest at PGE2 of 1 µg/mL (have not shown). Therefere, PGE2 1 µg/ml were used. The effect of HTS on T-cell proliferation following PGE2-induced suppression was evaluated in Jurkat cells with 1 µg/mL PGE2, a concentration that suppresses T-cell proliferation, and HTS at 20 mmol/L and 40 mmol/L above isotonicity was added at the same time as PGE2. After incubation for 24 hours at 37℃, the resultant T-cell proliferation was determined by the MTT cell proliferation assay (ATCC).
Enzyme-linked immunosorbent assay (ELISA) for MIF
MIF concentration in culture supernatants was measured by sandwich enzyme-linked immunosorbent assay. Briefly, 2 µg/mL of monoclonal capture antibody (R&D Systems, Minneapolis, MN, USA) was added to a 96-well plate and incubated for 2 hours at room temperature. After incubation, the plates were incubated in blocking solution comprised of PBS containing 1% bovine serum albumin (BSA) and 0.05% Tween 20 for 2 hours at room temperature. The test samples and standard recombinant MIF (R&D Systems) were added to the plates, and the plates were incubated for overnight at 4℃. The plates were then washed four times with PBS containing Tween 20, 200 ng/mL of biotinylated detection monoclonal antibodies (R&D Systems) was added, and the plates were incubated for 2 hours at room temperature. Subsequently, the plates were washed, streptavidin-alkaline-phosphatase (1:2,000; Sigma-Aldrich Co.) was added, and the reaction was allowed to proceed for 2 hours at room temperature. The plates were washed four times, and 1 mg/mL of p-nitrophenylphosphate dissolved in diethanolamine (Sigma-Aldrich Co.) was added to induce the color reaction, which was stopped by adding 50 µL of 1 N NaOH. The optical density at 405 nm was measured on an automated microplate reader (Bio-Rad Laboratories Inc., Hercules, CA, USA). A standard curve was generated by plotting optical density versus the log of the MIF concentration. The experiments were conducted 10 times.
Western blot analysis for MIF expression
Expression of MIF protein was quantified by Western blot analysis. Proteins (20 µg/sample) were fractionated on an 15% sodium dodecylsulfate-polyacrylamide gel (Bio-Rad Laboratories Inc.) and transferred onto a nitrocellulose membrane. Membranes were blocked for 1 hour in 5% BSA (Sigma-Aldrich Co.), and then incubated with a primary antibody, anti-human MIF (1:250; R&D systems). After washing, membranes were incubated with 1:2000 horseradish peroxidase-labeled goat anti-mouse antibody (R&D systems) as the secondary antibody. The proteins were detected using SuperSignal (Thermo Fisher Scientific Inc., Rockford, IL, USA) chemiluminescence kit.
Statistical analysis
One-way analysis of variance was performed to evaluate the significance of difference between the experimental groups. For a single comparison of two groups, a Student's t-test was used with SPSS ver. 12.0 (SPSS Inc., Chicago, IL, USA). Data are expressed as mean ± SD. Values of P < 0.05 were considered significant. All experiments were performed in triplicate.
RESULTS
The effect of HTS on PGE2-induced Jurkat cell suppression
Jurkat cells were plated on 96-well culture plates at a concentration of 2.5 × 106 cell/mL in cell culture media. PGE2 at 1 µg/mL inhibited Jurkat cell proliferation by 14.7% (P < 0.05) (Fig. 1). The addition of HTS restored Jurkat cell proliferation suppressed by PGE2 to control levels, as measured using an MTT cell proliferation assay (P < 0.05). There was no statistical difference in restoration by either HTS20 or HTS40 (Fig. 1).
The effect of HTS on MIF concentrations in the cell supernatant
To determine the relation between HTS and MIF on Jurkat cell proliferation, MIF levels were measured in the cell supernatant. The MIF level was decreased by 0.98 ng/mL ± 0.38 in the supernatant of PGE2 stimulated cells, when compared to control levels (1.19 ng/mL ± 0.48). On the other hand, HTS20 restored and increased the MIF level (1.09 ng/mL ± 0.38) in PGE2-suppressed Jurkat cells (P < 0.05). PGE2-suppressed Jurkat cells treated with HTS40 had a higher MIF level (1.16 ng/mL ± 0.33) than HTS20-treated cells, however, there was not a statistically significant difference of MIF levels in the supernatant of HTS20 or HTS40-treated cells (Fig. 2).
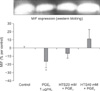
The effect of HTS on MIF expression
To determine the HTS effect on MIF expression, western blot analysis was performed. Correlating with the ELISA, levels of MIF protein expression were lower in PGE2-stimulating cells (16% decrease in band density) (P < 0.05). The addition of HTS to PGE2-stimulated cells increased MIF protein expression, with the highest expression in the HTS40-treated cells, however, there was not a statistically significant difference of MIF protein expression in the HTS20 or HTS40-treated cells (Fig. 3).
DISCUSSION
Trauma-induced suppression of cellular immune function likely contributes to sepsis, MODS, and death. This suppression is secondary to several immunomodulating factors including PGE2, transforming growth factor beta (TGFβ), interleukin 4 (IL-4), and IL-10, which block certain intracellular signaling events in T-cells [1]. T-cell dysfunction after traumatic stress is characterized by a decrease in T-cell proliferation, an aberrant cytokine profile, decreased T-cell monocyte interactions, and attenuated expression of the T-cell Receptor Complex [9-11]. The exact mechanism involved in the development of T-cell dysfunction is unknown. Under stressful conditions, macrophage are triggered to rapidly produce and release PGE2, a powerful endogenous immune suppressant [12]. PGE2 interacts with the corresponding membrane receptor of T cells, blocking T cell function by interfering with IL-2 gene expression at multiple stages [13-15]. In addition, PGE2 is a potent activator of cAMP-dependent protein kinase, which regulates expression of p27kip1 and cyclin D3 to suppress proliferation of leukemic T cell lines, including Jurkat cells, by arresting cells in the G1 phase of the cell cycle [15]. Therefore, the experiments were performed with PGE2, as it has been shown that PGE2, at a concentration of 1 µg/mL, causes inhibition of T cell proliferation by about 15%. Moreover, HTS restored suppressed PGE2-induced T cell proliferation. Hyperosmolality may play an important role in the HTS-enhancement of T-cell proliferation and hypertonicity increased IL-2 expression in T-cell proliferation [16-18]. However, the mechanism explaining the HTS-mediated restoration of T cell function is still unclear.
MIF may serve as a general marker for systemic inflammation in septic and non-septic acute critical illness [5-7]. Furthermore, MIF is a cytokine that is secreted by the anterior pituitary and immune cells in response to surgical stress, injury, and sepsis. This cytokine appears to be a critical regulator of the inflammatory pathways, leading to systemic inflammatory response syndrome and subsequent MOSF [4,19]. Besides, activated T cells produce MIF, and neutralizing anti-MIF antibodies inhibit T-cell proliferation and IL-2 production. T cells also release MIF in response to glucocorticoid stimulation and MIF acts to override glucocorticoid inhibition of T-cell proliferation, as well as interleukin 2 and interferon γ production [3]. And MIF also reduces pro-oxidative stress-induced apoptosis and nitric oxide induced apoptosis. As to a contribution by MIF to the regulation of cell survival, the inhibitory effect of MIF on apoptosis could correlate with its reported stimulatory effects on cell proliferation, even though cellular apoptosis and cell proliferation do not simply oppose each other [20-22].
That is to say, MIF exhibits pro- and anti-inflammatory activities, and regulates cell proliferation and survival. Therefore, we know that T cell immune restoration by HTS occurs, at least in part, by a MIF-mediated mechanism. In our experiment, PGE2 inhibited Jurkat cell proliferation and decreased MIF levels, when compared to control levels. The addition of HTS increased MIF production, as compared with PGE2-stimulated Jurkat cells in concordance with restored PGE2-suppressed Jurkat cell proliferation. Considering side effect of HTS, HTS at sodium concentrations of 160 mM showed sufficient result in this experiment. It has been suggested that HTS could be a proliferation regulator by a MIF-mediated mechanism. In order words, PGE2 reduces expression of cyclin D and induces expression of p27kip1, while MIF regulates the expression of the cell cycle regulators cyclin and p27 [15,23,24]. Therefore, HTS could restore PGE2-suppressed Jurkat cell proliferation through MIF production. Cho et al. [25] showed that NF-kB inhibition leads to increased synthesis and secretion of MIF in human CD4+ T cells. Notably, HTS also inhibits NF-κB, as compared to isotonic saline, in our experiment (unpublished results). However, the mechanism of MIF regulation of HTS is not yet explained.
Unfortunately, the present study is associated with certain limitation. First, PGE2 were used alone as immunosuppressant in present study. If another immuno-suppressant (IL-4, IL-10, et al.) were used, the present study might have more interesting outcome. Second, This study was done about MIF only, so other cytokine were not used. And ELISA and western blot analysis were performed for MIF levels. However, if another cytokine and another method for MIF levels (realtime PCR on the expression of mRNA for MIF) are used, the present study might have more interesting outcome, therefore, we will consider conducting further experiments by using other cytokine and methods in the future.
The role of HTS in restoring Jurkat cell proliferation suppressed by PGE2, at least in part, should be mediated through a MIF pathway. Further studies are needed to determine the effects of HTS and MIF.


 XML Download
XML Download