

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Menopause is induced by the decrease of ovarian hormonal secretion, especially estradiol. Women experience it usually around their late-forties, but can happen at any age. Various physiological changes due to menopause start gradually before the definite menopause. Menopause is natural course, but still the symptoms of it bother middle-aged women physically and emotionally. The most common symptoms are hot flush, headache, weight gain, vaginal dryness, depressive mood, agitation, forgetfulness and loss of libido.
Recently there have been many studies about depression of middle-aged women, reflecting increasing concern about it. It has been years since hormone replacement therapy (HRT) is applied to relive post-menopausal symptoms, but there are still controversies about its utility due to its side effects. However, there have been many studies which prove its effects to prevent osteoporosis and cardiovascular disease and to relieve those postmenopausal symptoms.1,2 Furthermore, some investigators emphasize the role of estrogen receptor in postmenopausal women, based on the increased expression of estrogen receptors in vaginal cells after administrating estrogen.3 Omega-3 fatty acid is one of naturally existing polyunsaturated fatty acids (PUFA) which have 18 to 24 carbons and 3 to 6 double bonds between carbons, one of which exists at the third carbon atom from the end of the carbon chain. It is one of the essential fatty acids, meaning that they cannot be synthesized by the human body. However, human have limited ability to synthesize it when they have ingested shorter form of omega-3 fatty acids, alpha-linolenic acid (ALA), usually found from plant oils. Human can transform ALA to the longer form, eicosapentaenoic acid (EPA), and then to the most crucial form, docosahexaenoic acid (DHA), even with less efficiency. The converting rate is not high, which is 5% for men, and little higher for women. Omega-3 fatty acids are expected to be effective substitutes to be a treatment for various postmenopausal symptoms. Erythrocyte levels of n-3 PUFA were positively correlated with bone mass.4
Ezrin/radixin/moesin (ERM) merlin family consist of 4.1 band superfamily, which has important role to control cell morphologic structure by connecting cell membrane and actin after phosphorylation.5,6,7 Activated ERM protein directly binds to actin filaments. Ezrin is an important protein to maintain cytoskeleton, which control the interaction between cell membrane and cytoskeleton.8 Several studies have found that estrogen activates the ezrin.9,10
We previously studied about the ezrin, vitamin D receptor (VDR), estrogen receptor in vagina after cowpea formula groups in ovariectomy mouse model.11 We thought dietary pattern is correlated with cytoskeleton factors in vagina. We planned to find out the effect of estrogen or omega-3 fatty acids on the expression of ezrin protein in the female vaginal cells.
There has been no study about how dietary intake may influence the ezrin protein except our previous data. We consider this study will be the first research to analyze the relationship between dietary intake of omega-3 fatty acids, estrogen level, and ezrin protein activation in the vaginal cells.
Materials and Methods
1. Postmenopausal rat model and diet
After getting approval of animal lab institutional review board (IRB) of the institution, three-week-old female Wistar/ST rats were raised after one week of adjustment period. The rats were then divided into three groups, for three different kinds of diet; general diet, 1% omega-3 fatty acid diet, and 2% omega-3 fatty acid diet. After eight weeks of having assigned diet, the nine-week-old rats had oophorectomy under general anesthesia. One centimeter of skin cut was made on the both sides of the back, and the abdominal muscles and fascia were cut until ovaries are exposed. After excision of the ovaries, abdominal fascia, muscles and skin were closed using silk and nylon suture.
2. Subcutaneous estrogen injection
One week of convalescence period is given after the oophorectomy, with the same diet previously they had. Each dietary groups are then divided into two subgroups; one subgroup for subcutaneous estrogen injection, the other for subcutaneous omega-3 fatty acids injection. Twelve weeks from the start of assigned diet, the rats are sacrificed.
3. Tissue preparation
Finally, according to the diet and injection type, rats were divided into 6 groups (Table 1). For each group, 4 rats were assigned. After 12 weeks from the start of assigned diet, the rats were sacrificed and their vaginas were harvested. Each vagina was divided into two pieces (one for Western Blotting, the other for paraffin embedding). For further study, one piece was stored in -80℃ until Western Blot analysis and the other piece were fixed in 4% paraformaldehyde for 48 hr. Then, fixed tissues were dehydrated through a series of graded ethanol baths to displace the water, and then infiltrated with wax. The infiltrated tissues were then embedded into paraffin blocks. For staining, tissues were cut into 5 mm thick sections using a mi crotome. Five-micron tissue sections were collected on poly-L-lysine-coated slides (Sigma-Aldrich Corp., St. Louis, MO, USA). Each tissue section was deparaffinized in xylene and rehydrated through a graded series of ethanol. Paraffin sections from each specimen were stained with hematoxylin and eosin to examine the full-thickness vagina.
To figure out ezrin and merlin, immunofluorescence staining and Western Blot analysis were performed.
4. Immunofluorescence
Paraffin section slides were washed in phosphate-buffered saline (PBS; pH 7.4) three times for 5 minutes. For antigen retrieval, slides were heated in a microwave oven at 600℃ with 0.01 M sodium citrate buffer (pH 6.0) for 10 minutes. After 1 hr cooling at room temperature, the slides were washed three times for 5 minutes in PBS.
Tissue sections were covered with ice-cold 100% methanol (use enough to cover completely to a depth 3-5 mm) and incubated in methanol for 10 min at -20℃, followed by washing three times with PBS. The slides were then incubated in a humidified chamber with blocking buffer (5% normal goat serum + 0.3% Triton X-100 in PBS) for 1 hr at room temperature. Primary antibody was applied in a moist chamber overnight at 4℃. The primary antibodies used were rabbit anti-NF2 (merlin) polyclonal antibody (sc-331, 1:200 dilution, Santa Cruz Biotechnology, Santa Cruz, CA, USA), rabbit anti-ezrin (phosphor T567) polyclonal antibody (ab47293, 1:100, Abcam Inc., Cambridge, MA, USA), and mouse anti-ezrin monoclonal antibody (ab4069, 1:200, Abcam Inc., Cambridge, MA, USA). After three additional rinsing steps with PBS for 5 minutes, anti-rabbit IgG H&L (Alexa Fluor® 647 conjugated) antibody (ab150079, 1:250, Abcam Inc., Cambridge, MA, USA) and anti-mouse IgG H&L (Alexa Fluor® 488 conjugated) antibody (ab150113, 1:250, Abcam Inc., Cambridge, MA, USA) was added separately for 1 hr at room temperature in dark. After being washed with PBS, coverslip slides with fluoroshield mounting medium with 4,6-diamidino-2-phenylindole (DAPI; ab104139, Abcam Inc., Cambridge, MA, USA). All experiments were executed with control staining without the primary antibody to ensure that negative controls remained unstained.
5. Western blot analysis
Vaginal tissues were snap freeze in liquid nitrogen. Freezing tissues were ground in liquid nitrogen into a powder with a mortar and a pestle. Lysis buffer consisting of 15 mM sodium chloride, 1.0% Triton X-100, 0.5% sodium deoxycholate, 0.1% sodium dodecyl sulfate (SDS), 50 mM Tris (pH 8.0), 1 mM ethylendiaminetetraacetic acid (EDTA) Protease Inhibitor Cocktail (Roche, Mannheim, Germany), and Phosphatase Inhibitor Cocktail 2, 3 (Sigma-Aldrich Corp., St. Louis, MO, USA) was added in the powder and the mixture was transfer into an eppendorf and incubated with agitation for 30 min at 4℃.
After the insoluble fractions were removed by centrifugation at 13,000 rpm for 20 min at 4℃, the supernatants were collected and protein concentration was determined by detergent compatible (DC) protein assay (Bio-Rad Laboratories, Berkeley, CA, USA). The protein lysate were separated by 10 or 12% SDS-polyacrylamide gel electrophoresis (PAGE) and transferred electrophoretically on to polyvinylidene difluoride (PVDF) membrane. Blots were blocked with 5% (w/v) skimmed milk in TBS-T (TBS with 0.1% Tween 20 [v/v]) for 1 hr at room temperature. After 3 times washing with TBS-T, blots were incubated at 4℃ temperatures overnight with a mouse monoclonal IgG1 anti-ezrin (ab4069, 1:1000, Abcam Inc., Cambridge, MA, USA), a rabbit polyclonal IG anti-Thr567 phospho-ezrin (ab47293, 1:1000, Abcam Inc., Cambridge, MA, USA), and rabbit anti-NF2 (merlin) polyclonal antibody (sc-331, 1:200 dilution, Santa Cruz Biotechnology, Santa Cruz, CA, USA). The membrane were than washed three times with TBS-T and incubated for 1 hr with anti-rabbit or mouse IG-conjugated with horseradish peroxidase. Enhanced chemiluminescence detection of Western Blot was performed by using the Western Blotting Luminol Reagent (sc-2048 Santa Cruz Biotechnology, Santa Cruz, CA, USA), according to the manufacturer's protocol.
Results
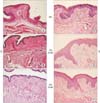
1. Effect of estrogen and dietary DHA on vaginal epithelial morphology
The atrophic vagina epithelium of ovariectomized rats was 2-3 cell layers thick (Fig. 1A-1C). In response to estrogen, vaginal epithelium are proliferated and differentiated. Vagina of estrogen injection group showed thicker and well differentiated features including cornification and keratinization (Fig. 1D-1F).
However, there were no significant responses to omega-3 fatty acid. Although omega-3 fatty acid composition changed in diet, vaginal epithelial morphology unchanged. Omega-3 fatty acid could not induce proliferation in the vaginal epithelium when compared with the effect of estrogen.
2. Ezrin and merlin expression on vagina
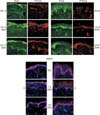
Ezrin, phosphorylated ezrin and merlin definitely expressed in basal layer. Because vaginas of oil injection group were atrophied, they had only basal layer in vagina (Fig. 1A-1C). However, in estrogen injection group, their vaginal epithelium showed full layers (from basal to apical layer) (Fig. 1D-1F).
Ezrin, phosphorylated ezrin and merlin expressed mainly in basal and parabasal layer. Their expressions were primarily cytoplasmic and strongest in epithelium. Additionally, the lamina propria showed focal expression of ezrin and phosphorylated ezrin. However, merlin did not express in lamina propria (Fig. 2).
In Western Blot analysis, we could present expression of ezrin and merlin in vagina in each group. Among dietary groups, there were no significantly differences in ezrin and merlin expression. Omega-3 fatty acid composition in diet did not affect expression of ezrin and merlin in rat vagina (Fig. 3).
However, estrogen presented significant impact on expression of ezrin and merlin. When comparing estrogen injection group with oil injection group, ezrin and merlin were more expressed in estrogen injection group than oil injection group. Phosphorylated ezrin showed similar pattern of ezrin. It supposed that almost of expressed ezrin would be activated form, phosphorylated ezrin.
Discussion
We confirmed the ezrin and merlin expression in vagina difference after estrogen at oophorectomy state mice. Ezrin and merlin are influenced by estrogen after menopause, but little influenced by omega-3 fatty. We would like to reveal association ezrin and merlin expression in vagina and omega-3 fatty.
The importance of omega-3 fatty acids for growth and health of human has been noticed, and various studies have followed to prove it. Proven effects so far are to prevent cardiovascular disease, stroke, osteoporosis, and cancers. Especially in obstetrical area, there have been several reports about increased incidence of having premature birth when pregnant women have not enough omega-3 fatty acids. It is recommended for women who are planning for pregnancy to take enough omega-3 fatty acids for its ability to help formation of new tissues in fetus, especially like neural development.12 However, omega-3 fatty acids has little effect in vagina.
At these days, as the age of women at the menopause has increased, the rate of intake of enough nutrients like omega-3 fatty acids, omega-6 fatty acids, dietary fiber, vitamin A, carotene, vitamin D, vitamin E, thiamine, riboflavin, pyridoxine, niacin, folic acids, vitamin C, calcium, phosphate, sodium, potassium, and iron is reported to be significantly low.13 Omega-3 fatty acids and gamma-linolenic acid like evening promise oil are available on the market, and these dietary supplements seem to qualify certain standards since these are analyzed by gas chromatography (GC)-flame ionized detector (FID).14
Ezrin protein is complicated tubular structure which has important roles in vaginal cells such as to support the structure or to participate in reproductive function. We have previously studied ezrin's location and its activation in twenty healthy women's vaginal cells under light microscope, electron microscope, and immunohistochemical stain.5 According to the results, ezrin protein is distinctly aggregated around membrane, and focally localized around cellular junction.5 It is expressed more when estrogen level is high, and on the keratinized epithelial cell. Based on those findings, we could conclude the protein has a role to control the interaction between vaginal cells, elasticity of the vaginal tissue and vaginal condition in a response to estrogen and other external environments.5
Other recent studies found out ezrin also takes an important role in metastasis of cancer and aggravation of several cancers such as rhabdomyosarcoma, pancreatic cancer, or ovarian cancer.6 Especially in ovarian cancer, in vitro studies showed the cellular proliferation level was directly proportional to the level of expression of ezrin protein, which could support a possibility of ezrin protein as a parameter to show malignancy of ovarian cancer.6
Despite of these various ongoing studies, there has been no study about how dietary intake may influence the ezrin protein. We consider this study will be the first research to analyze the relationship between dietary intake of omega-3 fatty acids, estrogen level, and ezrin protein activation in the vaginal cells.
We try to figure out the effect of omega-3 fatty acids on the vagina of postmenopausal women and its utility as a treatment for postmenopausal symptoms, and then to emphasize the importance of the dietary management after menopause.


 XML Download
XML Download