

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Preeclampsia is one of the most common obstetrical complications affecting up to 8% of all gestations1 and remains major causes of maternal and neonatal mortality and morbidity in Korea and worldwide. It typically presents after 20th week of gestation and is characterized by new-onset hypertension with proteinuria and/or edema during pregnancy.2 It is accepted that abnormal implantation of the blastocyst and abnormal placental bed remodeling or other causes of reduced placental perfusion are prerequisites for preeclampsia,1 but the mechanistic links between this step and the development of preeclampsia are still largely unknown.
A number of molecular pathways have been suggested to be involved in the cascade from placental events to maternal dysfunction. One of them which has been the subject of intense research is the production of the soluble form of the vascular endothelial growth factor (VEGF) receptor-1. The soluble VEGF receptor-1 is also denoted as soluble fms-like tyrosine kinase-1 (sFlt-1) as it belongs to fms-related tyrosine kinase family.3 The proto-oncogene fms encodes the tyrosine kinase transmembrane receptor. It has been shown that sFlt-1 is produced in the placenta during preeclampsia and is found to be elevated in the circulation of preeclamptic women, often before the onset of maternal symptoms.3,4 In addition, numerous experimental models have demonstrated the implication of sFlt-1 overexpression in the development of preeclampsia.5,6,7,8 In maternal circulation, sFlt-1 binds to free VEGF and placental growth factor (PlGF), sequestering and making them unavailable for proper signaling.3
Advanced glycation end-products (AGEs) represent the molecular complexes generated as a result of non-enzymatic reactions of carbohydrates and oxidized lipid with proteins. These reactions leads to irreversible cross-linking of proteins, and as a consequence, loss of protein structure and function.9 Accumulation of AGEs contribute to diverse metabolic disorders such as diabetes,10 obesity,11 metabolic syndromes,12 and polycystic ovary syndrome.13 The women with such disorders, when in pregnancy, have a higher risk of preeclampsia.1 Previous studies demonstrated that serum AGEs in preeclamptic women were significantly higher than in healthy women.14,15 Accumulation of AGEs may induce oxidative injury and vascular perturbation in placental bed, leading to preeclampsia.16 A recent study demonstrated that AGEs might contribute to the pathogenesis of preeclampsia by promoting sFlt-1 production in extravillous trophoblasts, implicating AGEs in a novel bridge between metabolic disorders and preeclampsia.17
Sildenafil, a specific inhibitor of cyclic guanosine monophosphate (cGMP)-specific phosphodiesterase type 5 (PDE-5), has evolved from a potential anti-angina drug into an oral treatment for erectile dysfunction (Viagra®; Pfizer, New York, NY, USA), and more recently, into a new orally active treatment for pulmonary arterial hypertension (Revatio®; Pfizer, New York, NY, USA).18 Sildenafil prolongs the relaxation and vasodilatation actions of cGMP on corpus cavernosal smooth muscle cells by inhibiting the breakdown of nitric oxide (NO)-driven cGMP.18
Theoretically, a therapeutic approach that could cause dilatation of the uterine or systemic vessels could restore placental perfusion, vascular tones of systemic arteries and consequently ameliorate the preeclamptic symptoms. Considering the well recognized action of sildenafil as a vasodilating agent, it would not be absurd to expect beneficial effects from sildenafil to alleviate preeclamptic signs and symptoms. Although trials in human patients did not provide confirmative results, these studies have left open the possibility that PDE-5 inhibitors could be beneficial to the preeclamptic patient.19,20,21 Animal studies with rat models of preeclampsia induced by NG-nitro-L-arginine methyl ester (L-NAME) treatment22 and placental ischemia23 showed that sildenafil treatment significantly reduced the preeclamptic signs and symptoms, such as hypertension, proteinuria and fetal mortality. Furthermore, sildenafil was shown to suppress elevation of the plasma levels of anti-angiogenic factors including sFlt-1.24 These results raised hope for the therapeutic potential of sildenafil as an effective option for the management of preeclampsia.
JEG-3, a choriocarcinoma cell line, may serve as an appropriate in vitro trophoblastic model to study preventive and therapeutic agents in the field of preeclampsia.25 This could help to understand of the association between sFlt-1 and other gynecologic conditions such as endometriosis and menopause.26 These cells have been shown to preserve responses to hypoxia to increase sFlt-1 mRNA expression and protein release.27 In this study, we examined the effect of sildenafil on sFlt-1 release in the AGEs-treated JEG-3 cells, and evaluated the role of the heme oxygenase-1 (HO-1) as a key molecule to mediate the action of sildenafil.
Materials and Methods
1. Cell culture
JEG-3 choriocarcinoma cells obtained from American Type Culture Collection were routinely maintained on plastic culture flasks in Dulbecco's Modified Eagle's Medium/Nutrient Mixture F-12 Ham (DMEM/F12) supplemented with 10% fetal bovine serum (FBS), 50 IU/mL penicillin G and 50 µg/mL streptomycin. Cells were trypsinized when cells became confluent (approximately every 4-5 days) using 0.05% trypsin/0.53 mM ethylenediaminetetraacetic acid (EDTA) solution and re-seeded at one-sixth the original density. Before 1 h to exposure to experimental protocols, cells were switched to serum-free media.
2. Preparation of AGEs-BSA
AGEs-bovine serum albumin (AGEs-BSA) was prepared as previously described.28 Endotoxin-free BSA (Calbiochem, La Jolla, CA, USA) of 5 g was incubated with 9 g D-glucose in 100 mL phosphate buffer (0.2 M, pH 7.4). The mixed solution was filtered through 0.22 µm microporous membrane filter and then maintained in sterile conditions in the dark at 37℃ for 3 months. At the end of incubation, unincorporated glucoses and low molecular reactants were removed by dialysis overnight against 0.1 M phosphate buffer. For BSA control BSA was incubated under the same conditions without glucose. Every preparation was tested for endotoxin by the limulus amebocytelysate assay (E-Toxate kit; Sigma-Aldrich, St Louis, MO, USA), and stored at 4℃ for use when no endotoxin was detected.
The efficiency of AGE-BSA formation was assessed by fluorescence spectrophotometric analysis (excitation 360 nm, emission 450 nm) and measurement of Nε-carboxymethyl-lysine (CML) by enzyme-linked immunosorbent assay (ELISA). AGEs-BSA was characterized by a 16 fold increase in glycation fluorescence and a 86-fold higher CML concentration compared to the unmodified BSA (17.2 nmoL/mg protein in AGEs-BSA vs. 0.2 nmoL/mg protein in unmodified BSA).
3. Reverse transcription-polymerase chain reaction (RT-PCR)
Total RNA was extracted using Trizol (Invitrogen, Carlsbad, CA, USA). In order to remove contaminating genomic DNA, 1 mg of RNA was treated with DNase (DNase I Amplification Grade; Invitrogen, Carlsbad, CA, USA) at room temperature for 15 min and the reaction was terminated with 25 mM EDTA for 10 min at 65℃. RNA was quantified by spectrometry and used for quantitative RT-PCR (qRT-PCR). qRT-PCR assays were carried out to quantify mRNAs for sFlt-1, receptors for AGEs (RAGE), HO-1 and β-actin. Real-time quantitation was based on the LightCycler assay, using a fluorogenic SYBR Green I re action mixture for PCR with the LightCycler Instrument (Roche Diagnostics, Mannheim, Germany). The PCR reactions were carried out in a total volume of 20 µL, which included SYBR Green I with a Taq DNA polymerase reaction buffer, deoxynucleotide triphosphate mixture, 10 mM MgCl2, PCR-grade water, 2 µL of template DNA, and primers. The sequences of the chosen primers were given in Table 1.
4. ELISA
sFlt-1 protein was measured by ELISA by using Quantikine Human ELISA kit (R&D Systems, Minneapolis, MN, USA).
5. Analysis of intracellular reactive oxygen species (ROS) generation
The intracellular generation of ROS was measured using 2',7'-dichlorodihydrofluorescein diacetate (DCFH-DA). The nonfluorescent ester penetrates into the cells and is hydrolyzed to DCFH by the cellular esterases. The probe (DCFH) is rapidly oxidized to the highly fluorescent compound 2',7'-dichlorofluorescein (DCF) in the presence of cellular peroxidase and ROS such as hydrogen peroxide or fatty acid peroxides. Cells cultured in 24-well plate were preincubated in the culture medium with 10 M DCFH-DA for 1 h at 37℃. After the preincubation, cells were exposed to experimental protocols for various durations. Changes in DCF fluorescence was assayed using FACSort Becton Dickinson Flow Cytometer (Becton-Dickinson Bioscience, San Jose, CA, USA) and data were analyzed with CELLQuest Software.
6. Reporter gene assay
Luciferase reporter gene assay was performed to measure activity. Transient transfections were performed with Lipofectamine Plus Reagent (Invitrogen, Carlsbad, Ca, USA) using pNF-κB-Luc and pCMV-β-Gal plasmids (Clontech Laboratories Inc., Palo Alto, CA, USA) The reporter gene assay was performed according to the manufacturer-provided method.
7. Western blot analysis
HO-1 protein was determined by Western blot analysis. Total intracellular protein was extracted by 5X repeated freeze-thaw lysis in FT buffer (600 mM KCl, 20 mM Tris-Cl, pH 7.8, 20% glycerol, 0.4 mg/mL Pefabloc, 10 µg/mL leupeptin, 10 µg/mL pepstatin, and 5 µg/mL aprotinin). Protein concentration was determined by Bradford assay (Bio-Rad). For Western blots, 30 µg of protein was subjected to sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) on 4-0% gradient SDS-PAGE (Bio-Rad). Membranes were blocked with Odyssey blocking buffer (LI-COR, Lincoln, NE, USA) for 2 h at room temperature. Primary incubation was undertaken overnight at 4℃ with a rabbit anti-HO-1 polyclonal antibody (StressGen, San Diego, Ca, USA) at 1 : 2000 and a mouse anti-β-actin antibody (Gentest) at 1 : 5000. Secondary antibody incubation was done with Alexa Fluor 680 goat anti-rabbit (Molecular Probes) and IRDye 800 goat antimouse IgG (Rockland) for 1 h at room temperature. Fluorescence was detected on an Odyssey infrared imager (LI-COR) for simultaneous detection of both species. Blot analysis was performed with the supplied Odyssey software, and HO-1 was normalized to β-actin.
8. Transfection with HO-1 siRNA
siRNA and scrambled siRNA against human HO-1 were purchased from Santa Cruz Biotechnology (Santa Cruz, Ca, USA), and transient transfections were performed using the Superfect® transfection reagent from QIAGEN (Hilden, Germany). The cells were incubated with the siRNA and scrambled siRNA at a concentration of 100 nM for 16 h in serum free media.
Results
1. Stimulation of sFlt-1 mRNA expression and protein release by AGEs-BSA
We examined first the JEG-3 choriocarcinoma cells used in this study preserve the basal expression and release of sFlt-1. qRT-PCR and ELISA analyses showed that the JEG-3 cells preserve baseline sFlt-1 mRNA expression and protein release. Control BSA did not affect the baseline sFlt-1 mRNA expression and protein release, whereas AGEs-BSA stimulated both the sFlt-1 mRNA expression and protein release in a concentration-dependent manner. When cells were treated with 200 µg/mL AGEs-BSA for 24 h, there were 3.5 fold and 2.7 fold increases, respectively, in mRNA expression and protein release (Fig. 1).
To examine whether the effect of AGEs-BSA in this study resulted from interaction with RAGE, the effect of anti-RAGE antibody was determined. In the presence of anti-RAGE antibody, AGEs-BSA failed to increase the sFlt-1 mRNA expression and protein release, indicating that interaction with RAGE is essential for the process of cellular actions of AGEs-BSA (Fig. 2).
2. Effect of AGEs-BSA on ROS production and NF-κB activity
Increased ROS production and NF-κB activation has been shown to be implicated in the AGE-related pathophysiological processes.29,30 In significantly increased ROS production and NF-κB activity. Anti-RAGE antibody inhibited the AGEs-BSA-induced stimulation of ROS production and NF-κB activity (Fig. 3).
3. Inhibition of AGEs-BSA-induced cellular effects by sildenafil citrate
To determine the effect of sildenafil citrate on AGEs-BSA-induced cellular effects, cells were incubated with AGEs-BSA for 24 h in the presence of sildenafil citrate (indicated concentrations or 20 µM, unless otherwise indicated). Sildenafil citrate did not affect the baseline sFlt-1 mRNA expression and protein release (data not shown), whereas it inhibited the AGEs-BSA-stimulated sFlt-1 mRNA expression (Fig. 4A) and protein release (Fig. 4B) in a concentration-dependent manner. Sildenafil citrate also suppressed the AGEs-BSA-induced stimulation of ROS production (Fig. 5A) and NFkB activity (Fig. 5B). AGEs-induced cellular effects have been known to be closely related with increased expression of RAGE. In the present study, in cells treated with AGEs-BSA a significant increase in the expression of RAGE mRNA was observed. In these cells, as well as in BSA control cells, sildenafil citrate did not show any significant effect on the RAGE mRNA expression.
4. Upregulation of HO-1 expression by sildenafil citrate
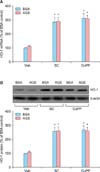
Several investigators showed that increased expression of HO-1 might inhibit sFlt-1 release and preeclamptic complications.31,32,33 Hence, we determined whether sildenafil citrate affects the HO-1 expression. qRT-PCR and Western blot analyses showed that sildenafil citrate significantly stimulated expression of HO-1 mRNA and protein in the cells treated with BSA control and AGEs-BSA. Treatment with AGEs-BSA alone did not affect the HO-1 mRNA and protein expression (Fig. 6).
5. Effects of HO-1 related molecules
In order to determine whether the effect of sildenafil citrate to inhibit sFlt-1 release and ROS production in cells treated with AGEs-BSA is related with the promoted expression of HO-1, we determined the effects of HO-1 related molecules. The HO-1 inducer, cobalt protoporphyrin (CoPP),34 and the metabolic product of HO-1, bilirubin,34 inhibited the AGEs-BSA-induced stimulation of sFlt-1 release. These effects were comparable to the sildenafil citrate-mediated inhibition of sFlt-1 release. On the contrary, the HO-1 inhibitor, zinc protoporphyrin IX (ZnPP IX)34 attenuated the effect of sildenafil citrate (Fig. 7A). Similar results were observed in the experiments to examine the effects on ROS production (Fig. 7B). These results strongly suggest that sildenafil citrate-induced inhibition of sFlt-1 release and ROS production in cells treated with AGEs-BSA is closely related with upregulated expression of HO-1.
6. Effect of HO-1 siRNA transfection
Small interfering RNA approaches were used to confirm further the role of HO-1 in the sildenafil citrate-induced inhibition of sFlt-1 release and ROS production in cells treated with AGEs-BSA. Western blot analysis showed that the basal expression of the HO-1 was sinificantly diminished by transfection with the HO-1 siRNA. In these cells, CoPP and sildenafil citrate failed to increase the expression of HO-1 (Fig. 8A). Likewise, in cells transfected with the HO-1 siRNA, sildenafil citrate failed to inhibit the AGEs-BSA-induced stimulation of sFlt-1 release and ROS production (Fig. 8B, 8C). These results confirm our hypothesis that upregulation of the HO-1 expression is the crucial mechanism to mediate the sildenafil citrate-induced inhibition of the sFlt-1 release and ROS production in cells treated with AGEs-BSA.
Discussion
Preeclampsia remains a major complication of pregnancy, affecting at least one out of twenty pregnancies.1 Besides the risk for mother and fetus during pregnancy and delivery, preeclampsia also impacts long-term health. Systemic review and meta-analysis reported that women with preeclampsia are at two-fold increased risk of developing cardiovascular disorders in later life.35 The later impact of preeclampsia is not confined to maternal health. It confers increased risk for cardiovascular complications to the offspring as well.36 Though the etiological cause of preeclampsia is not fully understood yet, diverse metabolic disorders have been recognized as predisposing causes toward the disease.1 AGEs have been implicated in the molecular bridge between the metabolic disorder and preeclampsia. Previous studies demonstrated that elevation of serum AGEs is closely related to the development of preeclampsia.14,15 A recent study demonstrated that AGEs might contribute to the pathogenesis of preeclampsia by promoting sFlt-1 production in extravillous trophoblasts, suggesting that AGEs might be an upstream molecule that promotes sFLt-1 expression in trophoblasts and contributes to the pathogenesis of preeclampsia.17 In the present study we also observed that AGEs-BSA increased sFlt-1 mRNA expression and protein release (Fig. 1). Although JEG-3 cells used in this study is a cell line established from choriocarcinoma, it has been shown to preserve trophoblastic cell-like characteristics, and might serve as an appropriate in vitro trophoblastic model for studies in various fields including preeclampsia.25 Thus, the results in this study can give us a valuable insight into trophoblastic mechanism responsible for the pathophysiology of preeclampsia.
AGEs exert their cellular effects through the interaction with RAGE, AGE-R3 (galectin-3), and maybe other AGE-binding proteins. Among them, RAGE is the best characterized AGE receptor and responsible for most of the deleterious effects of AGEs, whereas AGE-R3 is probably implicated in the removal of AGEs.37 In the presence of anti-RAGE antibody, AGEs-BSA did not affect the sFlt-1 mRNA expression and protein release, indicating that AGEs-RAGE interaction is essential for the process of cellular actions of AGEs-BSA (Fig. 2). AGEs-BSA-induced increases in ROS production and NF-κB activity are in accord with the previous understanding that increased ROS production and NF-κB activation is implicated in the AGE-related pathophysiological processes.29,30 In the presence of anti-RAGE antibody, AGEs-BSA failed to stimulate ROS production and NF-κB activity suggesting that these processes are also mediated through AGEs-RAGE interaction (Fig. 3).
Management of preeclampsia is still restricted by a lack of effective therapeutic agents, and there has been interest in using PDE-5 inhibitors in this patient population. Studies with several animal models of preeclampsia have demonstrated beneficial improvements in fetal outcome when treated with sildenafil.22,23,38,39 Herraiz et al.38 and Ramesar et al.22,24 demonstrated that the deleterious effect of NO synthas (NOS) inhibition during pregnancy could be significantly attenuated by sildenafil administration. Decrease in the bioavailability of NO is one of the most commonly recognized mechanisms of vascular dysfunction in preeclampsia. Normal pregnancy is associated with an increase in endothelium-derived NO, which is concomitant with overall increases in vasodilation and vascular compliance, which seems to be regulated by increased NOS activity.40,41,42 Indeed, pregnancy induces a significant increase in circulating levels of cGMP in the mother, suggesting increased NOS activity. Then, it would be logical to assume that a decrease in NO could be a major contributor to the endothelial dysfunction, and likewise, a restoration of NO bioavailability or cGMP with PDE-5 inhibitors could have a palliative effect in the preeclampsia patient. Further understanding of preeclampsia could improve general conditions of postpartum women and possibly connected to general welfare of menopausal women.
In the present study sildenafil citrate suppressed sFlt-1 mRNA expression and protein release in cells treated with AGEs-BSA in a dose-dependent manner (Fig. 4). Likewise, it inhibited the increase of ROS production and NFkB activity in these cells (Fig. 5A, 5B). These observations could provide us with a new insight into pharmacological action of sildenafil citrate. In addition to its well-recognized vasodilatory effect, the therapeutic targets of sildenafil citrate for preeclampsia may be widened to include the mechanisms within trophoblastic cell per se. In this regard, a previous report is notable, which has shown that systemic administration of sildenafil to L-NAME treated preeclampsia model rats decreased the plasma levels of anti-angiogenic factors including sFlt-1.24
Expression of RAGE proteins is known to be regulated by AGEs. In this study, increased expression of RAGE mRNA was observed in cells treated with AGEs-BSA indicating that AGEs-BSA upregulated the expression of RAGE. Sildenafil citrate did not affect the basal or AGEs-BSA stimulated expression of RAGE, indicating that action of sildenafil citrate does not seem to be related with regulation of RAGE expression.
One of the potential tools which has been proven effective in numerous experimental forms of hypertension is the manipulation of the HO-1 pathway.43,44,45,46 HO-1 produces two bioactive compounds as products of heme metabolism: carbon monoxide (CO) and bilirubin. Bilirubin functions as a powerful antioxidant.47,48 As production of ROS has been known to be a major component of varied forms of hypertension, moderate increase in bilirubin could act to decrease overall oxidative stress. CO acts as a potent vasodilator, functioning in a manner similar to NO.49 Of particular interest to preeclampsia, HO-1 and CO have been shown to directly inhibit production of VEGF and interferon-γ-induced secretion of sFlt-1.31
The present study strongly suggests the involvement of HO-1 in sildenafil citrate-induced inhibition of sFlt-1 release and ROS formation. qRT-PCR and Western blot analyses showed that sildenafil citrate upregulated the HO-1 expression (Fig. 6). Furthermore, the HO-1 inducer CoPP29 and metabolic product of HO-1 bilirubin29 also inhibited sFlt-1 release and ROS production in cells treated with AGEs-BSA, whereas the HO-1 inhibitor ZnPP IX29 antagonized the effect of sildenafil citrate (Fig. 7). Finally, small interfering RNA approaches were used to confirm further the role of HO-1 in the action of sildenafil citrate. In cells transfected with the HO-1 siRNA, sildenafil citrate failed to inhibit the sFlt-1 release and ROS production (Fig. 8). These results, taken together, strongly suggest that upregulation of the HO-1 expression is the crucial mechanism responsible for the sildenafil citrate-induced inhibition of the sFlt-1 release and ROS production in cells treated with AGEs-BSA.








 XML Download
XML Download