

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Acute lymphoblastic leukemia (ALL) has a high relapse rate, and chemo-resistance is a major characteristic of the disease [12]. One of the main factors affecting a clinical result in ALL is cellular drug sensitivity. Currently, glucocorticoids (GCs) are the most effective drugs to treat pediatric ALL. In the treatment of ALL, early response to prednisolone (a GC) is one of the most helpful prognostic factors [3]. Moreover, GC resistance in patients with ALL leads to treatment failure and consequent relapse [45]. Therefore, by targeting dominant markers of cancers, such as tumor cell resistance, chemo-resistance, and late relapses, survival of patients with cancer can be improved [6]. Deficiency or resistance to apoptosis are hallmarks of cancer and are caused by overexpression of anti-apoptotic proteins [7]. Overexpression of anti-apoptotic proteins such as BCL-2, BCLXL, and MCL-1 has been demonstrated in ALL cell lines. Moreover, the upregulation of BCL2 gene expression levels can help ALL cells to survive, even in the absence of optimal conditions for cell culture; for example, a lack of stromal derived growth factors [8910]. Therefore, an attractive approach to treat ALL is the inhibition of the anti-apoptotic/pro-survival members of the BCL-2 family of proteins. Resveratrol (3, 4, 5-trihydoxystilbene) is a member of the phytoalexins, which are found in many food products, such as grapes, berries, and peanuts. Several plants have been introduced as a source of resveratrol, and certain factors, such as UV irradiation, stress, and injury, can trigger the production of this compound [11]. Previous studies revealed certain interesting medical properties of resveratrol, including cardio-protective, anti-inflammatory, anti-oxidant, and growth-inhibitory activities [1213141516]. Resveratrol can overcome chemo-resistance in tumor cells. This property of resveratrol has been linked to controlling apoptotic pathways, modulating drug transporters, and downregulation of tumor cell proliferation proteins [17]. BCL2 family members, such as BCL-2, BCL-XL, MCL, BCL-W, AL/BFL-1, and BCL-B/BCL-2L10 are pro-survival factors that prevent tumor cells from undergoing apoptosis, while BAX, BAK, and BOK/MTD encode pro-apoptotic proteins that are responsible for inducing cell death [18]. Impairment of the pro-apoptotic and anti-apoptotic balance leads to disruption of apoptosis pathways in the affected cells. It has been widely proposed that frequent overexpression of BCL2 mRNA, combined with downregulation of BAX mRNA, in malignant conditions is one of the underlying mechanisms in pathogenesis of ALL [19]. In this study, we analyzed the effect of different doses of resveratrol (15, 50, and 100 µM), and prednisolone (700 µM), separately, and as a combined treatment, on the expression levels of BAX (pro-apoptotic) and BCL2 (anti-apoptotic) in the ALL cell line, CCRF-CEM. We also evaluated the effect of resveratrol and prednisolone on apoptosis in this cell line.
MATERIALS AND METHODS
Cell lines and culture conditions
T-ALL CCRF-CEM cells were purchased from Pasteur Institute of Iran (Tehran, Iran), and cultured at 4×105 cells per dish in Roswell Park Memorial Institute (RPMI) 1640 medium (GIBCO/Invitrogen, Grand Island, NY, USA) supplemented with 10% heat-inactivated fetal bovine serum (GIBCO), 50 µg/mL streptomycin, and 50 µg/mL penicillin, and maintained in a humidified incubator in 5% CO2 at 37℃.
Treatment
Resveratrol and prednisolone (98% purity, Sigma-Aldrich, Germany) were dissolved in ethanol. CCRF-CEM cells were treated separately and in combination with resveratrol (concentrations of 15, 50 and 100 µM) and prednisolone (700 µM). After 12, 24, and 48 hours, the cells were collected for RNA extraction.
RNA extraction and cDNA synthesis
Isolation of total RNA was performed using an RNX Plus kit (Cinnagen, Tehran, Iran) according to the manufacturer's protocol. Complementary DNA (cDNA) was synthesized from total RNA using a cDNA synthesis kit (Bioneer, Daejeon, Korea) following the manufacturer's instruction.
Quantitative real-time reverse transcription polymerase chain reaction (qRT-PCR)
The expression levels of BAX and BCL2 were determined using a SYBR Green qRT-PCR kit (Bioneer, Daejeon, Korea) according to the manufacturer's protocol. PCR reactions were performed using specific primers for BAX, BCL2, and the β-actin gene (ACTB) (Takapouzist, Tehran, Iran). The β-actin gene was used as the housekeeping gene to normalize the gene expression values. For graphing purposes, the relative gene expression in untreated cells, as a control, was set to 1. The following primers were used: BAX forward primer: 5′-GCCCTTTTGCTTCAGGGTTT-3′; BAX Reverse primer: 5′-TCCAATGTCCAGCCCATGAT-3′; BCL2 Forward primer: 5′-CGGAGGCTGGGATGCCTTTG-3′; BCL2 Reverse primer: 5′-TTTGGGGCAGGCATGTTGAC-3′; ACTB Forward primer: 5′-GAGACCTTCAACACCCCAGCC-3′; ACTB Revers primer: 5′-AGACGCAGGATGGCATGGG-3′. The reactions were carried out in triplicate using an Eco Real-Time PCR system (Illumina, San Diego, CA, USA).
Apoptosis analysis by flow cytometry
Cell death was assessed by Annexin V and Propidium Iodide (PI) staining (Exbio, Vestec, Czech Republic) according to the manufacturer's protocol. The cells were analyzed using a fluorescence activated cell sorting (FACS) caliber Flow cytometer (BD, San Jose, CA, USA) to determine the percentage of apoptosis.
Statistical analysis
The statistical calculations were performed using GraphPad Prism software (GraphPad Prism software Inc., La Jolla, CA, USA). The Kruskal-Wallis test and Dunn's test were used as multiple comparison tests to compare the statistical differences between more than two groups. P<0.05 was considered statistically significant.
RESULTS
The effect of resveratrol and prednisolone on the expression levels of BAX and BCL2
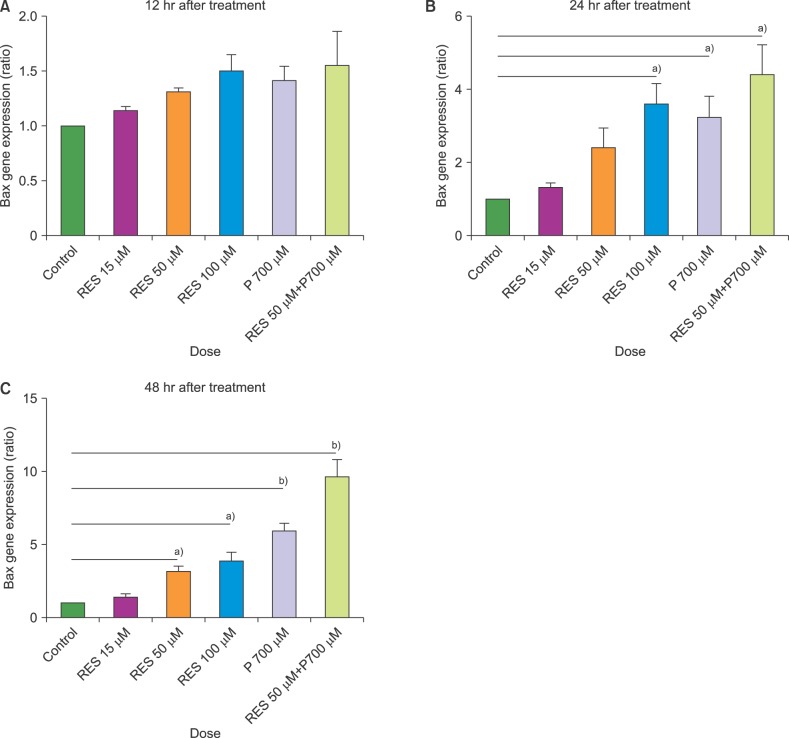
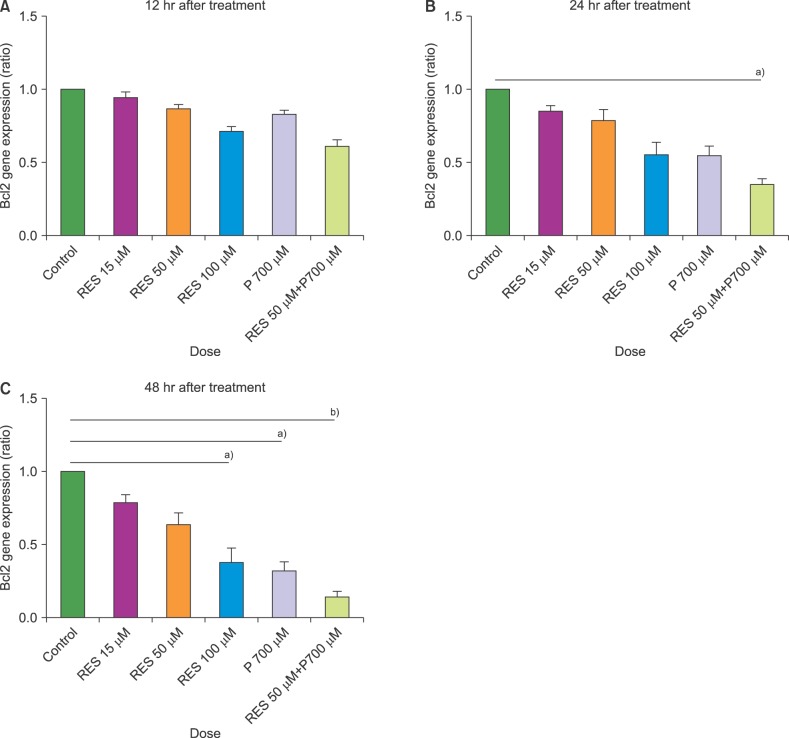
The expression levels of BAX and BCL2 were assessed in cultured T-ALL CCRF-CEM cell lines at 15, 50, and 100 µM concentrations of resveratrol and/or 700 µM of prednisolone versus untreated cells, using qRT PCR. No significant changes in the expression levels of BAX and BCL2 were observed at 12 hours. After 24 hours of treatment, the mRNA levels of BAX increased significantly under treatment with resveratrol at 100 µM, prednisolone at 700 µM, and a combination of resveratrol (50 µM) and prednisolone (700 µM). Only the combined treatment significantly decreased the mRNA levels of BCL2. The expression of BCL2 remained unchanged in the other conditions. Except for 15 µM resveratrol, we observed a significant increase in the mRNA level of BAX after 48 hours of treatment under all tested conditions. After 48 hours, a significant decrease in the expression of BCL2 was observed only using the combined treatment of resveratrol (RES) and prednisolone (PRE) at the following doses: 50 µM RES+700 µM PRE (Fig. 1, 2).
Apoptosis analysis by Flow cytometry
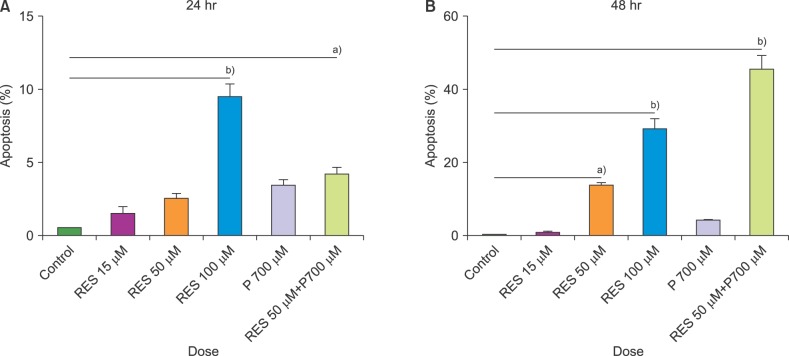
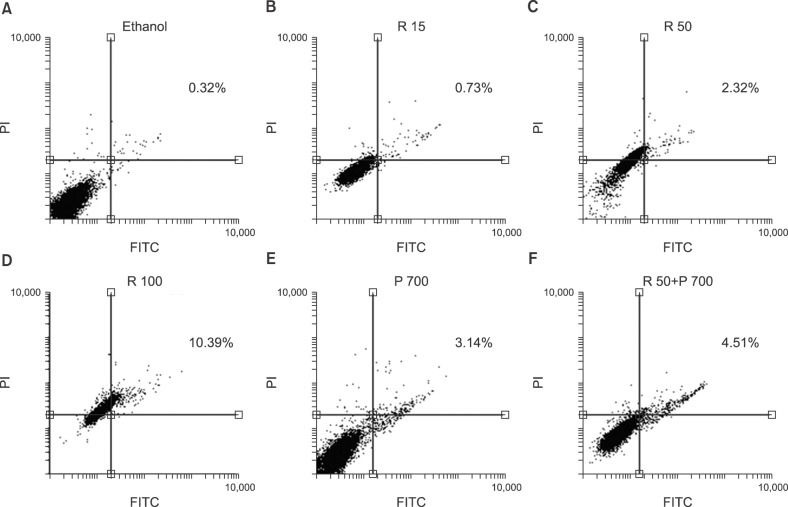
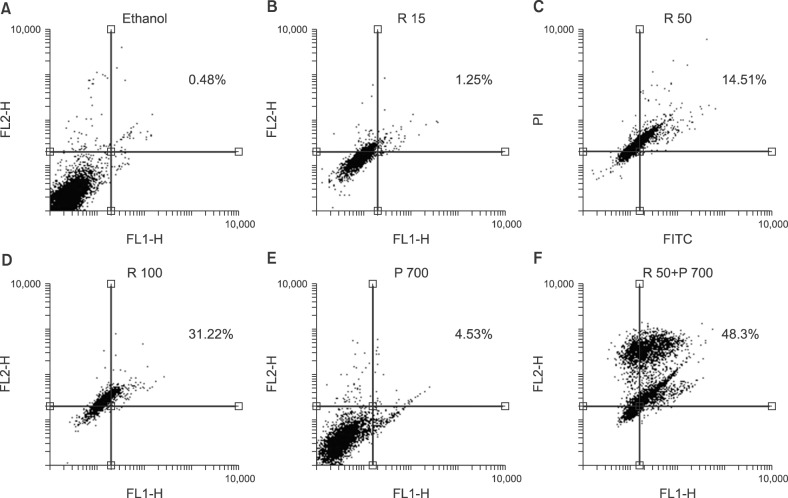
Apoptosis evaluation in CCRF-CEM cells treated with combined and separate treatment of resveratrol and prednisolone was performed using flow cytometry after 24 and 48 hours of treatment (Fig. 3, 4, 5). Ethanol-treated cells were used as the vehicle control. According to the obtained data, treatment with different doses of resveratrol and prednisolone significantly induced apoptosis in a time and dose-dependent manner in CCRF-CEM cells (Fig. 3A, B).
DISCUSSION
ALL is the most common pediatric cancer, which initiates from the bone marrow, followed by progressive invasion into other tissues [20]. Reduction of apoptosis or resistance to apoptotic pathways plays a critical role in pathogenesis of ALL. There are several ways to reduce or resist apoptosis in malignant cells. Consequently, to achieve optimal outcomes, stimulation of apoptosis is the therapeutic target of many drugs [20]. Knowledge of the molecular pathways that contribute to the development of leukemia might guide the development of new treatments to improve patient survival [21]. The overexpression of BCL2 in almost all types and subtypes of leukemia reveals the critical role of this protein in the pathogenesis and development of the disease. Although BCL2 is the most well-known member of the family, data indicate that other BCL2 family proteins, such as BAX, also play key roles [21]. In the present study, we examined the effect of resveratrol and prednisolone on BAX and BCL2 expression levels in ALL CCRF-CEM cells. Based on the qRT-PCR results, we observed that treatment time had a critical role in the effect of resveratrol; however, the effect of treatment time was not observed in cells treated with prednisolone. CCRF-CEM cells are a chemo-resistant cell line and perhaps for this reason, they did not show the expected apoptosis induction over the time course of the experiment. Currently, efforts to improve the survival of patients with cancer are based on approaches targeting chemotherapy resistance (chemo-resistance) and relapse, which are typical characteristics of ALL [22]. Resveratrol has diverse biological effects on cancerous cells, such as anti-proliferative, anti-inflammatory, and chemo protective activities [121314]. Some in vitro studies have demonstrated the potency of resveratrol to provoke apoptosis and inhibit the proliferation of a diversity of human cancer cell lines [23]. Recent research has described the chemo-prevention effect of resveratrol in combination with other chemotherapeutic drugs or cytotoxic factors in drug resistant tumor cells [13]. The anti-carcinogenic effect of resveratrol seems to be linked to its anti-oxidant property. It was revealed that this compound inhibits cyclooxygenase, hydroperoxidase, protein kinase C, BCL2 phosphorylation, Akt, a kinase binding position (focal adhesion kinase), nuclear factor-kappa B (NF-κB), matrix metalloprotease-9, and regulators of the cell cycle. Jazirehi and Bonavida [24] demonstrated the synergistic apoptotic effect of resveratrol and paclitaxel to selectively modify the expression levels of regulatory proteins in the apoptotic signaling pathway in resistant non-Hodgkin lymphoma and multiple myeloma cell lines. Fulda and Debatin [25] reported that resveratrol is an effective sensitizer for chemotherapeutic agents that induce apoptosis, such as doxorubicin, cytarabine (AraC), actinomycin D, paclitaxel (Taxol), and methotrexate, in a variety of human tumor cell lines via induction of cell cycle arrest and downregulation of Survivin expression. Bhardwaj et al. [26] showed that resveratrol could overcome chemo-resistance by downregulation of the NF-κB and signal transducer and activator of transcription 3 (STAT3) pathways. By inhibition of the NF-κB and STAT3 signaling pathways, resveratrol increases the release of BAX and the activation of caspase-3, which led to improvement of the apoptotic and anti-proliferative activities of Velcade and thalidomide in multiple myeloma cells. That study reported that resveratrol caused downregulation of various proliferative and anti-apoptotic gene products, such as Cyclin D1, cIAP-2, XIAP, Survivin, BCL-2, Bcl-XL, BFL-1/A1, and TRAF2. An elevated ratio of BAX/BCL2 was also reported previously in leukemia cell lines CEM-C7H2 and Jurkat, as a result of resveratrol treatment, suggesting that it exerts its pro-apoptotic effects by influencing intrinsic pathways [27]. Resveratrol induces apoptosis in B acute lymphoblastic leukemia, which is resistant to CD95-dependent signal transduction through the mitochondrial/caspase-9 pathway, as well as via the CD95-independent pathway [28]. Another study reported that resveratrol inhibited proliferation of two cell lines (OCMI2 and OCI/AML3) by arresting the cells in S phase [29]. By contrast, Cecchinato et al. [30] demonstrated that resveratrol upregulates the levels of the pro-apoptotic protein p53, inhibits the PI3K/Akt pathway, activates GSK-3β, and inhibits the NOTCH survival signaling pathways in MOLT-4 ALL cells. At a result, resveratrol induced apoptosis in a time- and dose-dependent manner in MOLT-4 cells by inhibiting the NOTCH signaling pathway and interfering with the p53 and PI3K/Akt pathways. Based on these previous studies, we aimed to combine resveratrol as a supplementary drug with prednisolone as a first line treatment. We did not achieve the expected results at 12 and 24 hours after treatment with the combined dose of resveratrol and prednisolone. However, after 48 hours, we observed that the combined treatment significantly increased the induction of apoptosis. The qRT-PCR result was in complete accord with the observed induction of apoptosis using the combined treatment. In the present study, we selected two doses of resveratrol (50 and 100 µM) to combine with prednisolone. Resveratrol at 15 µM was not combined with prednisolone because of its low effectiveness. One of the problems that we encountered was the percentage of cell necrosis that was observed in combined treatment of prednisolone and 100 µM of resveratrol. Hence, we used 50 µM of resveratrol as a selected and final dose to combine with prednisolone for gene expression assessment. Ultimately, our findings confirmed that resveratrol caused overexpression of BAX, decreased the expression of BCL2, and induced apoptosis in dose- and time-dependent manners via regulation of anti-apoptotic and pro-apoptotic proteins of the BCL-2 family. In addition, the flow cytometry results showed that resveratrol could induce apoptosis in a dose- and time-dependent manner in the CCRF-CEM cell line. In addition, treatment of CCRF-CEM cells with the combination of resveratrol and prednisolone confirmed the synergistic effect of these two compounds.
In conclusion, flow cytometry and RT-PCR assays showed that resveratrol could enhance the effects of a glucocorticoid drug (prednisolone) in the treatment of ALL.
 XML Download
XML Download