

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Mesenchymal stromal cells (MSCs) are spindle-shaped adherent cells of high plasticity that can be isolated from various sources, especially from the bone marrow, which is one of the most common tissues used for their isolation [1]. Owing to their potential for multilineage differentiation, secretion of important biomolecules, and immunomodulatory properties [2], MSCs are attractive candidates for cell therapy in treating immune disorders and in the field of regenerative medicine.
Although MSCs from the bone marrow are relatively easy to obtain, they are only available in limited amounts, accounting for around 0.01% of all nucleated bone marrow cells. Therefore, in vitro cell expansion is required before their usage for clinical applications. Expansion of MSCs has been typically conducted using traditional two-dimensional (2D) adherent culture conditions as a standard technique. Although the ease of this technique is its primary advantage, it has been demonstrated that the cells thus produced eventually lose their stemness, which is accompanied by replicative cell senescence, loss of differentiation potential, and reduced paracrine capability [34]. Studies with different culture systems have led to the concept that maintenance of progenitor properties generally requires conditions that best simulate a tissue-specific microenvironment, which is very difficult to achieve with a 2D culture system [5]. As such drawbacks currently limit the therapeutic potential of 2D-cultured MSCs, alternative culture methods, including three-dimensional (3D) culture, have been investigated to best preserve the cells' progenitor properties, and this has emerged as one of the most important topics in MSC research.
There is growing interest in culturing adherent cells using 3D techniques, which best recapitulate in vivo conditions [6]. 3D culture methods promote the potential of MSCs to differentiate to multiple lineages, consequently augmenting their tissue-regenerative and reparative effects [7]. Recent studies have also demonstrated that MSCs obtained from 3D culture have properties that could enhance their therapeutic potential [8].
The mechanisms contributing to the maintenance of the stemness and differentiation potential of MSCs have been under active investigation with the goal of identifying key regulators, which can be modified to enhance the therapeutic potential of MSCs. In addition to direct genetic modification of such regulators [9], control of gene expression as well as other genomic functions could be achieved by epigenetic modifications, for example by changing the local chromatin configuration or nuclear architecture [1011]. In line with these epigenetic mechanisms, our team previously reported that epigenetic agents such as hypomethylating agents or a histone deacetylation inhibitor could change the gene expression pattern and function of cells [1213], which suggests that such epigenetic modification could also be used to change the characteristics of MSCs.
Specifically, we hypothesized that the progenitor properties of 2D-cultured MSCs could be altered by pretreatment with azacitidine (AZA), and consequently analyzed the effect of such a treatment on the multilineage differentiation potential, characteristics, and cellular senescence of MSCs under 2D and 3D culture conditions.
Go to : 
MATERIALS AND METHODS
Preparation and characterization of MSCs
Adult C57BL/6 mice (7 weeks old) obtained from Orient Bio (Seongnam, Korea) were used in the experiments of this study with the approval of the Animal Experiment Board of the Catholic University of Korea. Bone marrow cells were collected by flushing the femurs and tibias of C57BL/6 mice with Dulbecco's modified Eagle medium (DMEM; Gibco, Carlsbad, CA, USA) supplemented with 1% antibiotics (10 U/mL penicillin, 10 g/L streptomycin; Gibco), and 10% fetal bovine serum (FBS). Cell immunophenotypes persistently positive for stem cell antigen-1 (SCA-1), CD44, and CD29, but negative for c-Kit, CD31, FLK-1, and CD34 were confirmed at passage 8 by flow cytometry (Becton Dickinson, San Jose, CA, USA). Cells obtained after passage 9 were used in all experiments. 3D cultured MSCs which was prepared according to the hanging-drop culture method. All experiments were performed in quadruplicate (N=4).
AZA treatment
MSCs were incubated with 1 µM AZA (Sigma-Aldrich, St. Louis, MO, USA) in conditional medium for 72 h. Controls consisted of MSCs cultured under the same conditions in the absence of AZA.
Cell proliferation assay
Cell proliferation, with and without AZA treatment, was analyzed after 1, 3, 5, and 7 days, using the colorimetric 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium (MTS) assay. At each time point, the cells were washed with phosphate buffered saline and incubated with 20% MTS in serum-free medium. After 4 h of incubation at 37℃ in a 5% CO2 incubator, aliquots were pipetted into a 96-well plate and the absorbance was measured in each well at 490 nm using a spectrophotometric plate reader (Molecular Devices, Sunnyvale, CA, USA).
Apoptosis assay
After harvesting, 2D-MSCs, AZA-treated 2D-MSCs, 3D-MSCs, and AZA-treated 3D-MSCs were stained with annexin V-fluorescein isothiocyanate (FITC) and propidium iodide (PI), and gated according to the ratio of PI negative/annexin V-FITC-positive cells for detecting the proportion of apoptotic cells. The analysis was performed with an LSR Fortessa flow cytometer (Becton Dickinson) using the FACs Diva 6 software.
Colony-forming unit-fibroblast (CFU-F) assay
MSCs were seeded in a culture plate at a density of 500 cells/well in growth medium, which was changed every 2–3 days. After 14 days of culture, the cells were stained with 1% crystal violet solution (Sigma-Aldrich, Gillingham, Dorset, UK) and the number of colonies (diameter≥2 mm) was counted.
Differentiation assays
Trypsinized MSCs were induced to undergo adipogenic and osteogenic differentiation. For adipogenesis, cells plated in 24-well plates and grown to confluence were incubated in adipogenic differentiation medium for 2 weeks (Thermo Fisher Scientific, Waltham, MA, USA), and were either used for RNA isolation for genetic analysis or stained with Oil Red O for marker detection (Sigma). For osteogenesis, cells grown on 24-well plates to confluence were incubated with osteogenic differentiation medium (DMEM with 10% MSC-FBS, 10−7 M dexamethasone, 10 mM β-glycerophosphate, and 0.3 mM ascorbic acid) [13] for 2 weeks, and were either processed for RNA isolation or stained with alkaline phosphatase (ALP) to highlight calcium deposits.
Real-time polymerase chain reaction (PCR) assay
Total RNA was extracted from MSCs using the TRIzol reagent (Invitrogen, Carlsbad, CA, USA). First-strand cDNA was prepared by reverse transcription with the Transcriptor reverse transcriptase (Roche, Mannheim, Germany) and the appropriate oligo (dT) primers, and stored at −20℃. Real-time PCR was performed using SYBR green (Roche) on a Roche QPCR system. The primer sets for the genes of Oct-4, Nanog, Cxcr4, Runx2, and Ppar-γ are listed in Table 1. Cycle threshold (Ct) values of Oct-4, Nanog, Cxcr4, Runx2, and Ppar-γ mRNA were normalized to that of β-2 microglobulin, which was used as an internal control. Relative expression levels were determined according to the ΔΔCt method.
Senescence assay
For evaluation of senescence markers, control and AZA-treated cells were stained with fresh senescence-associated β-galactosidase (SA-β-gal) chromogenic substrate solution (Cell Signaling Technology, Danvers, MA, USA), and imaged with a phase-inverted microscope (Leica Microsystem, Wetzlar, Germany).
Statistical analysis
All quantitative data were expressed as group means±standard deviations. For statistical analysis, the Mann-Whitney U test was performed using the SPSS software (SPSS, Chicago, IL, USA). P <0.05 was considered to denote statistical significance.
Go to :
RESULTS
Preparation of MSCs
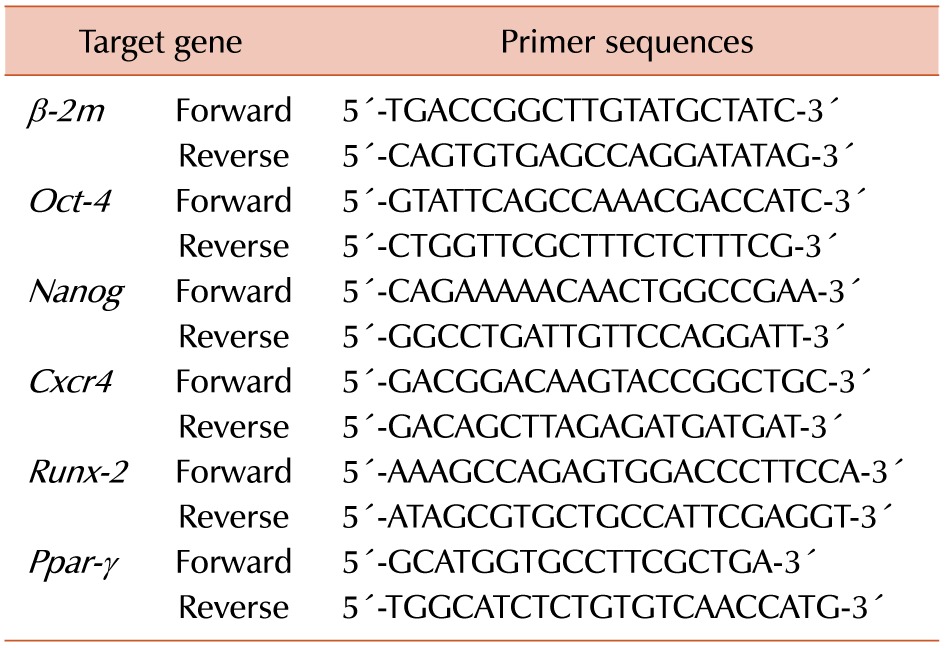
Phenotypic changes between 2D-MSCs, AZA-treated 2D-MSCs, 3D-MSCs, and AZA-treated 3D-MSCs were evaluated by flow cytometry. No changes in the MSC-negative markers CD31, CD34, c-Kit, and FLK-1 were detected across all cell groups. However, the expression of the MSC-positive markers CD44 and SCA-1 were significantly decreased in 3D-MSCs and AZA-treated 3D-MSCs than in the 2D-MSC groups (with or without AZA treatment). These results indicate that the 3D culture treatment induced a change in the MSC phenotype (Fig. 1A).
 | Fig. 1(A) Immunophenotyping results of mouse MSCs. After harvesting, 2D-MSCs, AZA-treated 2D-MSCs, 3D-MSCs, and AZA-treated 3D-MSCs were stained with antibodies and analyzed by flow cytometry. The cells were strongly positive for MSC-specific markers such as CD29, CD44, and SCA-1, and negative for the CD31, CD34, c-Kit, and FLK-1 markers. (B) Effects of AZA on MSC viability. Relative proliferation rates were determined by the MTS assay. (C) MSC apoptosis was determined by flow cytometry based on PI uptake and Annexin V-FITC labeling (N=4, a)P < 0.05, b)P <0.01).
|
Cell apoptosis and proliferation
Cell proliferation was assessed with the MTS assay. The results showed that AZA treatment did not influence the proliferation of either 2D-MSCs or 3D-MSCs. The proliferation rates of cells cultured in 2D were higher than those of cells cultured in 3D after 3 or 5 days of culture, whereas no significant difference was detected after 7 days of culture (Fig. 1B). However, both AZA-treated cell groups showed decreased levels of apoptosis, and apoptosis was lower in 3D-MSCs than in 2D-MSCs (Fig. 1C). These results demonstrate that cell proliferation and apoptosis were not affected by the culture conditions (N=4, a)P<0.05, b)P<0.01).
Osteogenic differentiation
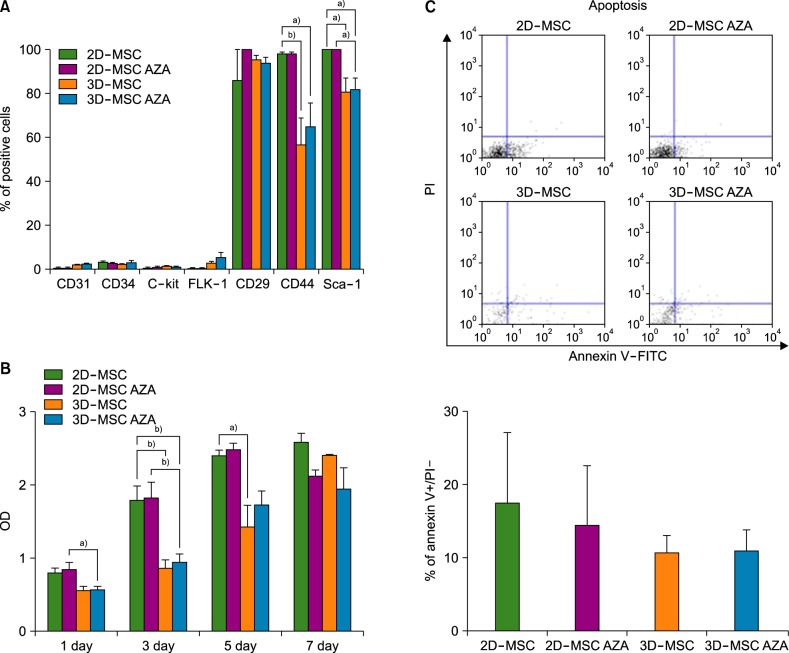
First, ALP staining was performed to identify the osteogenic potential of the different groups of cells. Osteogenesis was highest in AZA-treated 3D-MSCs, followed by that in 3D-MSCs, AZA-treated 2D-MSCs, and 2D-MSCs (Fig. 2A). Real-time PCR analysis revealed increased expression levels of the osteogenesis-related gene Runx-2 in 3D-MSCs than in 2D-MSCs, although the difference was not statistically significant (Fig. 2B). Furthermore, the Runx-2 expression levels were significantly increased in AZA-treated 3D-MSCs than in 2D-MSCs and AZA-treated 2D-MSC, but were not significantly different from those observed in 3D-MSCs. These results indicate that both AZA treatment and 3D culture could improve MSC osteogenesis (N=4, a)P<0.05).
Adipogenic differentiation
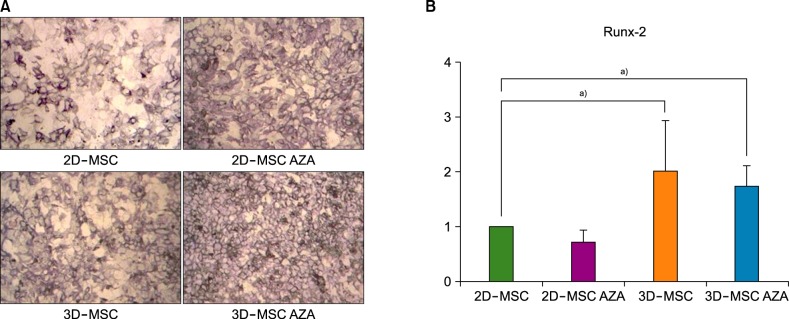
Adipogenesis, as identified with Oil Red O staining, was the highest in AZA-treated 3D-MSCs, followed by that in 3D-MSCs, AZA-treated 2D-MSCs, and 2D-MSCs (Fig. 3A). The real-time PCR results indicated increased expression levels of the adipogenesis-related gene Ppar-γ in AZA-treated 2D-MSCs, 3D-MSCs, and AZA-treated 3D-MSCs than in 2D-MSCs, although the differences were not statistically significant (Fig. 3B).
 | Fig. 3MSC adipogenesis. (A) Oil red O staining was performed after adipogenic differentiation of MSCs treated with or without AZA treatment. (B) The transcript levels of Ppar-γ were analyzed in the various MSC populations after adipogenesis induction with adipogenic medium for 72 h. β-2 microglobulin was used as an internal control (N=4).
|
Expression of colony-forming assay and stemness markers
To identify stemness changes resulting from AZA treatment in 2D- and 3D-MSCs, a CFU-F assay (Fig. 4A) and real-time PCR for the stemness-related genes Oct-4, Nanog, and Cxcr4 (Fig. 4B-D) were performed. As shown in Fig. 4, both assays demonstrated that neither AZA treatment nor 3D culture had an influence on MSC stemness.
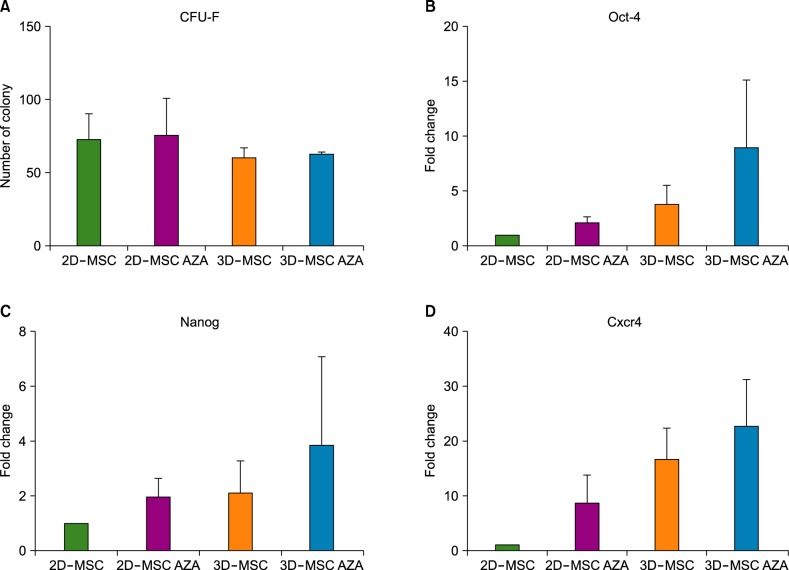 | Fig. 4CFU-F assay and expression of stemness markers. MSCs were plated at a density of 500 cells/plate and incubated for 14 days for the CFU-F assay. (A) The number of colonies (diameter≥2 mm) was counted. The expression levels of Oct4 (B), Nanog (C), and Cxcr4 (D) in control and AZA-treated MSCs were assessed by real time PCR analysis (N=4).
|
Effect of AZA treatment on MSC senescence
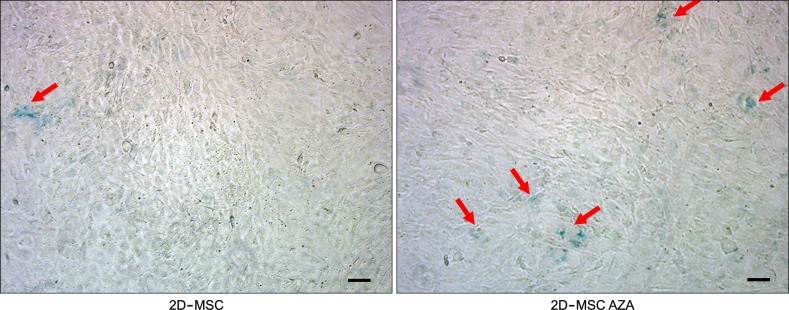
To identify any changes in MSC senescence induced by AZA treatment, an SA-β-gal assay was conducted with 2D-MSCs. A higher number of cells showed β-gal staining in the AZA-treated group than in the untreated group, indicating that AZA could induce MSC senescence (Fig. 5). The number of β-gal positive cells was 4-fold higher in AZA-treated cells than in 2D-MSCs (data not shown).
Go to :
DISCUSSION
Since their discovery in 1976 by Friedenstein et al. [14], MSCs have been mostly expanded in vitro on polystyrene tissue culture plates. The milieu of this conventional culture system is quite different from that of the tissue from which MSCs are extracted; 2D culture notably lacks the 3D architecture and composition of original tissues, as well as the oxygen tension, mechanical stimuli, and paracrine signaling from other cell types. Consequently, 2D expansion techniques tend to lead to the deterioration of the progenitor properties of MSCs [3415]. There have been many efforts to overcome these limitations by mimicking the natural cell environment, including the use of growth factors, reduction of oxygen tension, and development of 3D expansion systems [7161718]. Although traditionally expanded MSCs are still regularly used in clinical trials because of the relative ease of preparation, alternative expansion techniques that do not compromise the cells' differentiation properties need to be explored.
A key progenitor property of MSCs is their multilineage differentiation potential, which is the main advantage of their application in the field of cell therapy for tissue repair or regeneration. Given the importance of epigenetic mechanisms in stemness maintenance, differentiation, senescence, and transdifferentiation of MSCs, one potential expansion method that could retain the cells' differentiation power during long-term 2D expansion could be epigenetic cell modification [11]. For example, chromatin modification by epigenetic drugs has been proven to enhance multilineage differentiation of MSCs to osteocytes, adipocytes chondrocytes, and cardiomyocytes [192021]; however, the data regarding the influence of AZA on MSCs remain controversial [22]. We have previously observed that a combination of AZA and trichostatin A treatment enhanced multilineage differentiation, with AZA-induced hypomethylation being the main contributing mechanism [13]. However, in the present study, we observed no significant changes in the expression levels of key regulatory molecules in 2D-MSCs following AZA treatment. This discrepancy could be associated with the differences in the concentration of AZA used (1 µM in this study vs. 2 µM in the previous study) or the exposure and treatment time (induction for 3 days before differentiation in this study vs. 2 weeks during differentiation in the previous study). However, in the present study, 3D-MSCs showed enhanced differentiation potential, accompanied by activation of key molecules, and AZA treatment further enhanced their osteogenic differentiation. Rosca and Burlacu [22] also reported enhanced differentiation of AZA-pretreated MSCs and suggested that the subsequent optimal stimuli following AZA treatment may have contributed to this effect. Ultimately, the usefulness of AZA in the preparation of MSCs and its optimal dosage should be further explored to uncover the mechanisms underlying the observed results.
The mechanisms by which AZA enhances differentiation could be related to the basic action of the drug: removal of methylation and activation of transcriptionally silenced genes [1223]. Furthermore, a recent study showed that AZA converts somatic cells to multipotent stem cells [24], suggesting that the enhanced multilineage differentiation of MSCs observed could be a result of the AZA-induced activation of stemness-related genes that lead to enhanced progenitor properties. However, in this study, there were no significant changes in the transcript levels of Oct-4, Nanog, and Cxcr4, which are well-known genes involved in stemness maintenance. Moreover, we observed no differences in the number of colonies detected in the CFU-F assay among the different cell groups, further suggesting that AZA treatment did not affect the progenitor properties of MSCs. Of practical importance, AZA did not affect cell proliferation and apoptosis, although increased senescence has been observed [25]. These results require further investigation.
Collectively, our data suggest that 3D culture and AZA treatment could be used to enhance MSC differentiation. Nevertheless, further studies are needed to develop more efficient culture conditions and find the optimal AZA dosage that could maximize the positive effects of such culture conditions, especially when aiming to prepare MSCs with high osteogenic differentiation potential to enhance their therapeutic efficiency in regenerative medicine.
Go to :
 XML Download
XML Download