

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
The ABO gene, which is located on chromosome 9, has seven exons and is over 18 kilobases (kb) in length. The major coding region of ABO is found in exons 6 and 7 [1], and many ABO alleles have been described [2]. These alleles have minor differences in their genomic sequences, and most have emerged because of hybrid alleles, base insertions, deletions, or substitutions, or by splice site mutations [23]. Furthermore, the frequencies of these alleles differ among different populations and ethnicities. For example, the A101, O01, B101, A102, and O02 alleles are common in Asian populations [14]. Alleles such as O04 and A205 are found in the Han Chinese, and allele O06 is found in the Japanese [56]. Rare alleles such as cis-AB01, cis-AB02, and cis-AB06 are more frequently encountered in the Japanese, Korean, and Chinese populations [578].
The ABO locus has three main phenotypes, A, B, and O, with the combination of glucosyltransferases encoded by different alleles determining the A, B, AB, or O blood group phenotype [910]. Each phenotype can be distinguished serologically using anti-A and anti-B antisera generated by polyclonal or monoclonal techniques. These serological tests are important for matching the blood groups of acceptors and donors and not only enable the determination of additional subgroups, but also facilitate differentiation between weak A and B antigens [1112]. From an application perspective, allele and genotype studies of ABO are of primary importance in paternal discrepancy, forensics, organ transplantation, and population studies [11]. A number of strategies based on polymerase chain reaction (PCR) have been developed for ABO genotyping including restriction fragment-length polymorphism (RFLP) analysis and allele-specific PCR [13], single-strand conformation polymorphism analysis [14], amplified product length polymorphism (APLP), and sequencing [15]. To date, there has been no study in the western region of Saudi Arabia using these more current genotyping approaches. Therefore, the purpose of this study was to determine and evaluate the frequency and distribution of ABO phenotypes and their alleles using specific primers by multiplex PCR.
MATERIALS AND METHODS
Collection of blood samples
This prospective study was approved by the ethical committee of the Faculty of Medicine, King Abdulaziz University Hospital at Jeddah (Saudi Arabia). Peripheral blood was obtained with informed consent from 107 participants. ABO phenotypes were serologically determined with anti-A and anti-B antibodies using an AutoVue Innova (Ortho Clinical Diagnostics, Raritan, NJ, USA).
Genomic DNA extraction
Genomic deoxyribonucleic acid (DNA) from blood was extracted using the QIAamp DNA kit (Qiagen, Hilden, Germany). Briefly, 200 µL of sample was mixed with 20 µL of Qiagen protease in a microcentrifuge tube; 200 µL of buffer AL was added followed by thorough mixing and incubation at 56℃ for 10 min. Afterwards, 200 µL of absolute ethanol was added to the lysate, followed by gentle vortexing for proper mixing. Next, the mixture was transferred to a mini spin column for centrifugation and washed three time using buffer AWI before a final elution with 50 µL of AE buffer. Extracted DNA was quantified using a NanoDrop spectrophotometer (Thermo Fisher Scientific Inc., Wilmington, DE, USA). The method followed the procedure used by Muro et al. [16].
Multiplex PCR and agarose gel electrophoresis
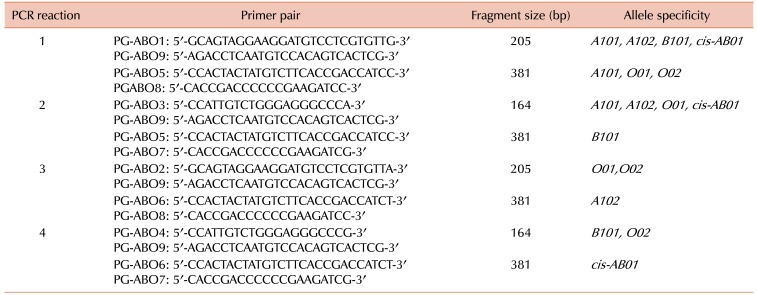
Primers used for PCR are listed in Table 1. The genotyping approach for this study was used previously in several studies [1718]. Four separate PCR reactions were performed using different sets of primers in each sample. Each 20 µL reaction contained 2 µL DNA mixed with 1 µL of each allele-specific primer (10 pmol), 10 µL master mix (2X), and 7 µL nuclease free water. PCR was performed in a thermal cycler (Applied Biosystems, Wilmington, DE, USA). The cycling conditions followed an initial denaturation at 95℃ for 3 min, followed by 35 cycles of denaturation at 95℃ for 40 s, annealing at 56℃ for 40 s, and elongation at 72℃ for 40 s, followed with a final elongation at 72℃ for 5 min. PCR products were separated by electrophoresis on 3% agarose gels containing ethidium bromide at 40 mV for 20 min followed by 70 mV for 30–40 min. A 100 base pair (bp) DNA ladder was used as a molecular marker (Invitrogen, Wilmington, DE, USA). The gel was visualized using a G-BOX UV transilluminator (Syngene, Cambridge, UK).
RESULTS
Phenotype distribution
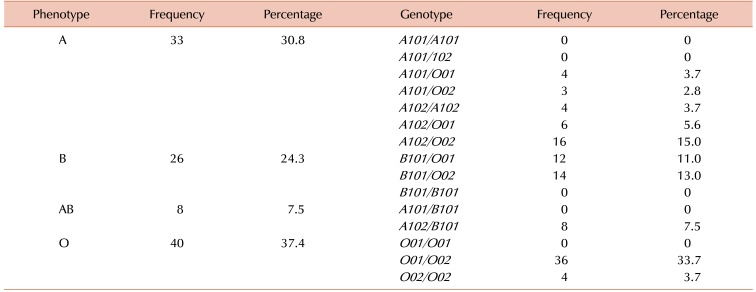
We found that serological ABO typing and phenotypes of study participants inferred from genotypes were concordant with an expected distribution between the A (N=33; 30.84%), B (N=26; 24.30%), AB (N=8; 7.48%), and O (N=40; 37.38%) blood groups.
Genotype distribution
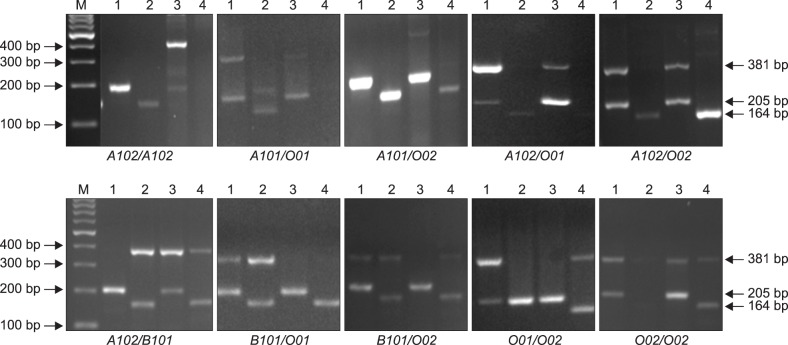
Out of 21 possible ABO genotypes, 10 genotypes were observed in our study group (Table 2). The O01/O02 genotype was the most frequent accounting for 33.6% of participants followed by the A102/O02 genotype that was found in 14.9% of participants. In addition, we found that frequencies of the B101/O02, B101/O01, A102/B101, and A102/O01 genotypes were 13.1%, 11.2%, 7.5%, and 5.6%, respectively. Genotypes such as A101/A101, A101/A102, A101/B101, O01/O01, and B101/B101 as well as rare genotypes including cis-AB01/O02, cis-AB01/O01, and cis-AB01/A102, were not be detected in our cohort. We found that eight individuals were homozygous for either A102/A102 (N=4) or O02/O02 (N=4), while the remaining 99 individuals were heterozygous.
Allele variation
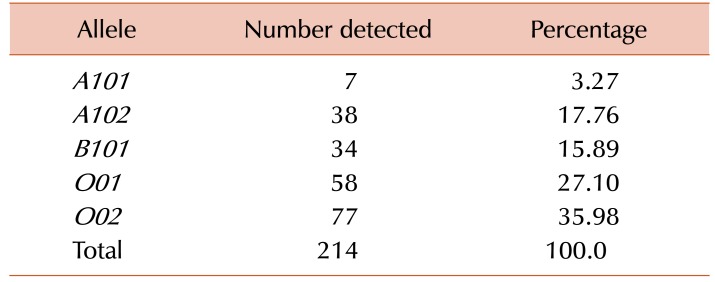
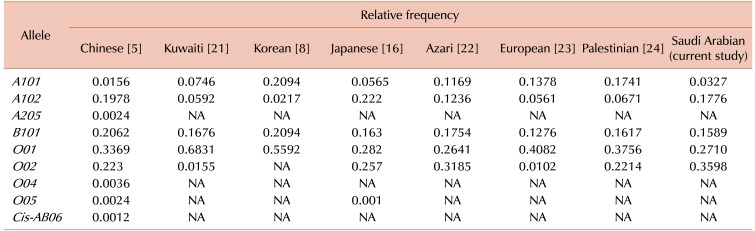
Primers used to amplify the six alleles of ABO are shown in Table 1. The frequency distributions of ABO phenotypes, genotypes, and alleles are summarized in Table 2 and Table 3. Our allele distribution consisted of A101 (N=7), A102 (N=38), B101 (N=34), O01 (N=58), and O02 (N=77). We found that O02 was the most abundant allele in our study cohort accounting for 36.0% of all alleles, while the A102 allele had a frequency of 17.8%. The frequencies of the B101 and O01 alleles were 15.9% and 27.1% respectively, and the A101 allele had the lowest frequency at 3.27%. We found that our western Saudi population had an increased frequency of the O02 allele compare to the O01 allele. Fig. 1 shows the electrophoretic patterns of the 10 recognized genotypes, while Table 4 provides a comparison of allele frequencies among different populations.
DISCUSSION
We found in our cohort from the western region of Saudi Arabia phenotype frequencies of 30.8%, 24.3%, 7.5%, and 37.4% for the A, B, AB, and O blood groups, respectively. Previous studies performed in other Saudi populations found different frequencies of 34.2%, 24.8%, 2.5%, and 38.5% for the A, B, AB, and O blood groups, respectively [19]. For all studies including the present one, blood group O has the highest frequency followed by A, B, and AB. This variation in ABO phenotype found between different Saudi populations is expected among ethnic groups [20]. For example, in the United Kingdom, blood group B has a low frequency (10%), whereas in India it is 18.8% [4].
We found 10 ABO genotypes in our cohort, of which O01/O02 was the most predominant found in 33.6% of individuals. These findings corroborate similar findings in a Han Chinese population where O01/O02 is the most frequently observed genotype (16.8%). In addition, we found that the frequency of the O02/O02 genotype in our Saudi population (3.7%) was comparable to that found in a Han Chinese population. Furthermore, and similar to our findings, another study did not find the A101/A101 genotype in their cohort [5]. However, our findings differed from those found in a Kuwaiti population where the homozygous O01/O01 genotype was detected in 47.0% of blood samples [21] whereas it has not been reported in a Saudi population [19]. In contrast, we found that the A102/O02 genotype was present in 14.9% of blood samples which was absent in the Kuwaiti population. Rare genotypes such as cis-AB01/O02, cis-AB01/O01, and cis-AB01/A102 were conspicuously absent in the current study with these latter alleles being detected among 13 different genotypes in the Korean population [8]. In addition, the cis-AB01 genotype has been reported in Japanese and Chinese populations [56]. The absence of these genotypes cis-AB01/O02, cis-AB01/O01 and cis-AB01/ A102 in studies from the Middle East indicates that the Middle Eastern population differs considerably from the Chinese, Korean, and Japanese populations in respect to these rare genotypes.
Of the six possible alleles, we found five alleles of which the most frequent allele was O02 with a frequency of 0.3598 followed by the O01 allele with a frequency of 0.271. A comparable study on ABO blood group genotypes in a Chinese population found that the O01 allele was the most frequent with a frequency of 0.3369 followed by the O02 allele. In addition, alleles A102 and B101 with frequencies of 0.1978 and 0.2062 in our cohort were comparable to frequencies of 0.1776 and 0.1589 respectively, in the Chinese cohort [5].
As anticipated, the frequency of various alleles in different populations around the world can differ widely because of ethnicity (demonstrated in Table 4). This variation has been widely observed in many studies [56821222324]. In the majority of populations examined, the A101 and A102 alleles are common, whereas the O02 allele is predominant in the Saudi population. In comparison, the O01 and O02 alleles are frequent in Han Chinese, Kuwaiti, and other populations with varying frequencies [521]. Similar differences in allele frequencies are found in the European, Japanese, German, Kuwaiti, and Indian populations [462123].
In conclusion, the present study reports the distribution of ABO phenotypes and genotypes in a group of individuals from the western region of Saudi Arabia. Using allele-specific multiplex PCR, 10 ABO genotypes were identified. In addition, this pilot study identified five alleles A101, A102, B101, O01, and O02, which were found at different frequencies in our cohort, and we performed a comparative assessment to other studies. Future plans include analyzing a larger number of samples to obtain a more comprehensive understanding of the ABO genotype pattern in Saudi Arabia.
 XML Download
XML Download