

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Maintenance of the homeostatic state is of prime importance in an organism and demands proper balance between cell death and cell division. Apoptosis, or programmed cell death, is a process that results in the controlled elimination of unhealthy or damaged cells. The phenomenon was first described by Kerr et al. in 1972 when they proposed a distinct 'cell deletion' pattern characterized by significant morphological changes [1]. Apoptosis is a complex cellular mechanism governed by several signaling molecules. There is a huge body of literature related to the significance of apoptosis in cell clearance and the maintenance of cell populations. Caspases, a group of cysteine proteases (cysteine aspartyl specific protease), are the key mediators of apoptosis [2]. Caspase-1, the first member in the caspase family, initially known as interleukin-1 beta-converting enzyme (ICE), was shown to have sequence similarity with Ced3 gene coding for CED-3 protein responsible for cell death in Caenorhabditis elegans suggesting their role in apoptosis [3]. The significance of caspases in physiology has led to intense research in this field. Caspases are synthesized as zymogens, possessing a pro-domain and a protease domain, and they are activated through allosteric conformational changes, proteolytic cleavage, or both [4]. Caspase family members are classified as upstream, or initiator, caspases (caspase-1, -2, -4, -5, -8,-9, -10, -11, and 12) and downstream, or effector, caspases (caspase-3, -6, -7, and -14) [5]. Caspases are also involved in non-apoptotic functions, including cell-cell communication [6], cytokine maturation [7], and inflammatory responses [8]. Recent studies, however, have introduced a new aspect to this process, citing their significance in cell development and differentiation. In some cell types, caspase-mediated apoptosis is mandatory for terminal differentiation. Caspase activation is also involved in the differentiation of erythrocytes [9], keratinocytes [10], skeletal muscle [11], lens fiber cells [12], as well as monocyte to macrophage differentiation [13]. Defective effector caspases, namely, caspase-3, hampers differentiation of some cell types, thereby suggesting a role for these apoptotic proteases in cell development.
Go to : 
APOPTOSIS
Apoptosis is a characteristic phenomenon undergone by every cell as a means to establish homeostasis in a cell population. This event, as reported by Kerr et al. is associated with significant distortions in cell morphology [1]. Apoptosis induced changes in cell morphology is initially marked by nuclear condensation [1]. Marked cells then undergo fragmentation, which results in the formation of apoptotic bodies in a process called blebbing. Apoptotic bodies prevent the leakage of immunogenic components (such as nucleic acids, oxidizing metabolites, and lysosomal enzymes) from the dying cells, thereby preventing the inflammation and autoimmune responses [14]. Apoptotic bodies are later phagocytosed by macrophages or other nearby cells in response to cell surface indicators of apoptosis, such as phosphatidyl serine or changes in carbohydrate moieties. [15]. Apoptosis is a complex, high coordinated process, involving several signaling molecules and other components, which eventually results in disposal of the damaged cell. Apoptosis occurs through either of the two different pathways: the intrinsic, mitochondrial pathway or the extrinsic, death receptor mediated pathway (Fig. 1).
 | Fig. 1Extrinsic and intrinsic pathways of apoptosis. The extrinsic pathway is initiated by ligation of death receptors with death ligands. This interaction results in the formation of the death inducing signaling complex (DISC), which contains the death receptor, an adaptor molecule, and pro-caspase 8. Adaptor molecules possess a death domain (DD) and a death effector domain (DED). At the DISC, caspase 8 is autocatalytically activated and subsequently transmits the death signal to effector executioner caspases, resulting in apoptotic cell death. The intrinsic pathway signaling cascade is triggered by a number of factors, including DNA damage, hypoxia, growth factor deprivation, and ER stress. The death signal is sensed initially by the BH3-only protein, which then interacts with the downstream mediators of apoptosis (BAX and BAK). BAX and BAK undergo distinct conformational changes, which lead to the formation of mitochondrial pores or increases the permeability of the mitochondrial outer membrane, thereby releasing apoptogenic compounds, e.g. cytochrome c. Released cytochrome c binds to APAF-1 to facilitate formation of the apoptosome, a wheel-shaped heptameric complex, which can then recruit and activate pro-caspase 9. As a consequence, caspase 9 activates effector caspases (caspase 3, 6, or 7) and eventually leading to apoptosis. Abbreviations: TNF, tumor necrosis factor; TRAIL, TNF related apoptosis inducing ligand; FADD, Fas-associated death domain protein; TRADD, TNF receptor-associated death domain protein; BID, BH3 interacting-domain death agonist; tBID, truncated BID; BAX, Bcl-2 homologous antagonist/killer; BAK, Bcl-2 associated X protein; APAF-1, apoptotic protease activating factor-1.
|
Go to :
EXTRINSIC AND INTRINSIC PATHWAYS OF APOPTOSIS
Execution of apoptosis is a regulated mechanism involving several components. The extrinsic and intrinsic pathways are the two most extensively studied apoptotic pathways. In the extrinsic pathway, caspase activation is initiated at membrane bound death receptor (DR) molecules that belong to the tumor necrosis factor (TNF) receptor superfamily [161718]. To date, eight death receptors have been characterized: tumor necrosis factor receptor-1 (TNFR1, also known as DR1), Fas (CD95/APO-1), TNF related apoptosis inducing ligand receptor-1 (TRAIL R1/DR4), TRAIL R2/DR5, DR3 (APO-3/TRAMP), DR6, nerve growth factor receptor (NGFR), and ectodysplasin A receptor (EDAR) [19]. Each of these have corresponding ligand molecules that belong to the TNF family of proteins, including TNF-α [20], Fas ligand (FasL/CD95L) [21], TNF related apoptosis inducing ligand (TRAIL) [22], and APO-3 ligand (APO-3L) [23]. The extrinsic pathway is invoked when death associated ligands bind to their respective death receptors. This ligation initiates a signaling pathway cascade that finally results in the execution of cell death [23]. Adaptor molecules possess a death domain and a death effector domain. The death domain mediates association with the death receptor; this association leads to the recruitment of the initiator caspase (caspase-8/caspase-10) to the ligand receptor complex. The adaptor molecule then mediates the interaction between the death receptor and the initiator caspase. This signaling complex, comprising the death receptor, the adaptor molecule, and the pro-caspase, is named the death inducing signaling complex (DISC) [24]. In the DISC complex, accumulation of pro-caspase-8 leads to its autocatalytic activation into caspase-8, which then transmits the death signal to the effector executioner caspases resulting in apoptotic cell death.
The intrinsic pathway of apoptosis is mediated by the release of effectors such as cytochrome C and SMAC/Diablo from the inter-membrane of mitochondria [25]. The release of such proteins into the cytosol triggers the signaling cascade that finally activates the executioner caspases, thereby resulting in cell death. DNA damage, growth factor deprivation, and endoplasmic reticulum (ER) stress act as common signals for the activation of this pathway, which is regulated by members of the Bcl-2 protein family [26]. Bcl-2 family members include both pro-apoptotic and anti-apoptotic factors. Pro-apoptotic factors include BAK, BID, BOK, BIM, BAD, NOXA, and PUMA; anti-apoptotic factors include Bcl-2, Bcl-XL, Bcl-W, MCL-1, and Bcl-B. The Bcl-2 proteins are further subdivided based on the identity and number of Bcl-2 homology (BH) domains they possess: 4 BH domains (BH 1-4) containing anti-apoptotic proteins; pro-apoptotic proteins with 3 BH domains (BH1-3) which comprise BAX, BAK, and BOK; and pro-apoptotic proteins with only the BH3 domain called BH3-only proteins (BID, BIM, BAD, and NOXA) [27]. Localization of these pro-apoptotic and anti-apoptotic proteins differ during normal growth conditions or in the presence of a death signal. Anti-apoptotic proteins are integral membrane proteins found in mitochondria [28], endoplasmic reticulum, or the nuclear membrane [29]. The death signal is sensed initially by the BH3-only protein, which then interacts with the downstream mediators of apoptosis (BAK and BAX) [30]. BAK and BAX undergo distinct conformational changes, which finally leads to the permeabilization of the outer mitochondrial membrane and the subsequent release of apoptogenic compounds such as cytochrome c. Released cytochrome c binds to apoptotic protease activating factor-1 (APAF-1) to facilitate formation of the apoptosome [25], a wheel-shaped heptameric complex (Fig. 1), which can then recruit and activate pro-caspase-9. As a consequence, caspase-9 activates executioner caspases (caspase -3, -6, -7), eventually leading to apoptosis.
Go to :
CASPASES IN HEMATOPOIESIS
Hematopoiesis is a remarkable process that results in the production of mature blood cells. Hematopoietic stem cells (HSCs) are the common progenitor cells from which different lineages of blood cells arise [31]. The process occurs as a result of finely orchestrated cell signaling networks that involve many factors, including cytokines and transcription factors. Hematopoiesis is strictly controlled by cell division, differentiation, and cell death, during which various caspases and apoptotic factors are involved (Fig. 2). Apoptosis is a major mechanism utilized by the hematopoietic system to regulate blood cell production according to the pathophysiological condition [32].
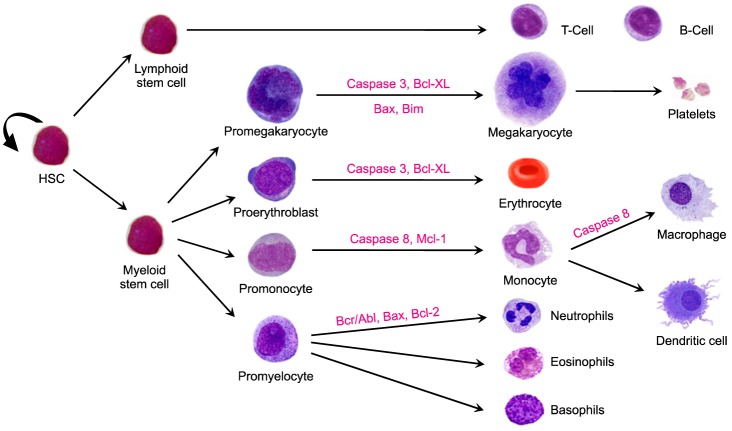
Caspases are evolutionarily conserved and present in almost every eukaryotic cell. The founding member of the caspase family, caspase-1, initially known as interleukin-1 beta-converting enzyme (ICE), was found to have a sequence similarity with CED-3, a protein in C. elegans that is responsible for cell death, suggesting that caspases may function similarly in mammalian cell death [13]. Since then, several caspases were identified. Caspases, in general, are synthesized as inactive zymogens with an N-terminal pro-domain region and a caspase catalytic region. Some caspases have a shorter pro-domain region while others possess a longer pro-domain region. Though these cysteine proteases are mostly associated with apoptotic cell death, some are also assigned non-apoptotic functions. Hematopoietic stem cells express CD95/Fas, which was shown by Barcena et al. to be expressed in primitive hematopoietic progenitor cells from human fetal liver [33]. Barcena et al. also found that Bcl-2 expression was higher in CD 95+ cells than in CD95- cells, suggesting the regulation of Bcl-2 expression during differentiation and a possible protective role against apoptotic signals [33]. Bcl-2, Bcl-XL, Mcl-1, and A1 are different members of the Bcl-2 family that are expressed on the surface of hematopoietic stem cells [34]. In a study conducted on erythroid progenitors, Bcl-XL was indispensable for the maturation of primitive and definitive progenitors [35]. Defect in Mcl-1 hampered the development of B- and T-lymphocytes and reduced the population of mature lymphocytes [36]. TRAIL, a death receptor molecule, has recently been found to have a non-apoptotic function, acting as both a promoter and as an inhibitor of maturation of erythrocytes, megakaryocytes, and monocytes [37]. SCL/Tal-1, a transcription factor, also exerts proliferative and anti-apoptotic functions in hematopoietic cells [21].
Go to :
APOPTOSIS IN ERYTHROCYTE DIFFERENTIATION
Erythrocytes, or the red blood cells, are produced in the bone marrow through a process known as erythropoiesis. Approximately 2×1011 erythrocytes are produced per day, and this estimation makes this population of blood cells the highest generated. The process involves several complex steps, from the differentiation of hematopoietic stem cells into multipotent progenitors to the commitment of the progenitors into erythroid progenitors as burst forming unit-erythroid (BFU-E) and colony forming unit-erythroid (CFU-E). CFU-E differentiate into proerythroblasts, which then form basophilic erythroblasts, the polychromatophilic erythroblasts, and, finally, the last nucleated cells of the erythrocyte progenitor acidophilic erythroblasts [38]. Acidophilic erythroblasts undergo enucleation, a characteristic feature of erythrocytes where the cell extrudes a nucleus and gives rise to reticulocytes that finally mature into erythrocyte. The major factors involved in the process are erythropoietin (EPO), which regulates the red cell number by controlling terminal differentiation, and GATA1, which governs their proliferation [39]. Erythroblast cells in their terminal differentiation stage express a very high level of Bcl-XL, an anti-apoptotic protein. Interestingly, primary erythroblasts expressed very little of the protein [40].
EPO promotes erythroid progenitor survival by repressing apoptosis. In the absence of EPO, Bcl-XL and Bcl-2 expression are down regulated [41] in the early stages of murine and human erythroblast differentiation, leading to activation of caspase-3 and ultimately to cell death [42]. GATA1, together with EPO, promotes cell survival by regulating Bcl-XL expression [43]. STAT5 is associated with the immediate-early induction of Bcl-XL through direct binding to the promoter of the Bcl-X gene [44]. However, towards the terminal stages of differentiation, transient expression of caspase-3 becomes indispensable. There are a significant number of studies that emphasize the use of apoptotic machinery during the later stages of erythropoiesis. The initial studies were carried out by Zermati et al. and addressed the role of caspases in erythrocyte differentiation [9]. They showed that the presence of z-VAD-fmk, a broad spectrum inhibitor of caspase, reduced terminal erythroid differentiation of CD36+ cells in a dose dependent manner. Caspase-3 was found to be transiently activated through the mitochondrial pathway. Interestingly, Zermati et al. also found that GATA 1, a substrate molecule for caspase-3, was spared although other substrates, such as acinus and lamin B, were cleaved. GATA 1 is associated with the chaperon molecule Hsp70, which protects it from caspase-3 mediated cleavage [45]. SCL/Tal-1, another transcription factor involved in erythropoiesis, is resistant to effector caspases and has been shown to be a negative regulator of erythroid differentiation [46].
Go to :
APOPTOSIS IN MEGAKARYOCYTE DIFFERENTIATION
Megakaryocytopoiesis is a tightly regulated, cell differentiation process that results in the formation of the most rare and intriguing population among the immune cells, the megakaryocytes. Megakaryocytes play a crucial role in maintaining the hemostatic condition, as they are the progenitors of platelets, clotting factors, which are critical when the immune response is compromised as the result of a tissue injury occurs. Megakaryocytes develop from hematopoietic stem cells under the influence of the cytokine, thrombopoietin (TPO), which specifically regulates maturation and platelet production [47]. In the absence of TPO, immature megakaryocytes undergo apoptosis [47]. The HSC forms the common myeloid progenitors and then forms megakaryocyte/erythrocyte progenitor (MEP), which gives rise to burst forming unit-erythroid (BFU-E) and high proliferative potential-colony forming unit-megakaryocyte (HPP-CFU-MK). HPP-CFU-MK then gives rise to BFU-MK from which CFU-MK develops. Immature megakaryocytes, the megakaryoblasts, arise from this CFU-MK which then differentiate into mature megakaryocytes, develop proplatelets, and finally shed platelets [48]. Megakaryocyte differentiation is characterized by the formation of long cytoplasmic projections termed proplatelets. Proplatelets undergo a drastic morphological change, which results in the conversion of these enormous cells into highly filamentous structure. These filaments further form node-like structures from which the mature anucleate platelets are shed. Because megakaryocytes are the rarest cell population in the bone marrow, in vivo studies of the development, differentiation and terminal differentiation of these cells into platelets have presented the field with many hurdles. Studies have shown that caspases are involved in some stages of development and differentiation of these cells.
Although development and differentiation of these cells have been extensively studied, a number of aspects are still ripe for research. One such aspect is the role of caspase in the processes. Several studies focusing on the role of caspase in the differentiation of megakaryocyte to proplatelets and then the subsequent platelet production have been conducted [4950]. Both megakaryocytes and platelets possess a functional apoptotic caspase cascade downstream of Bcl-2 family-mediated mitochondrial damage [51].
A study by de Botton et al. demonstrated the activation of caspase-3 in megakaryocytes just prior to the formation of proplatelets. Caspase-3 activation was localized in certain granular structures and a diffused activation of it in the cytoplasm lead to cell death. In the study, the authors found an intriguing phenomenon in which caspase-3, whose activation under normal conditions leads to apoptotic cell death, has spared the terminally differentiating megakaryocyte as there was no sign of DNA fragmentation, which is considered to be a characteristic indication for apoptosis in cells. Caspase-3 has non-apoptotic functions, including cell proliferation, cell cycle progression and cellular differentiation [52]. Inhibition of both caspase-3 and caspase-9 causes reduced proplatelet formation [49]. Mitochondrial membrane potential and membrane phosphatidyl serine asymmetry of proplatelet forming megakaryocytes were studied by Clarke et al. [50]. According to the study, the proplatelet extension of megakaryocytes, which exhibit compartmentalized caspase activation, retain these features whereas the main cell body showed apoptotic features, including nuclear condensation and caspase activation. Interestingly, ligation of the membrane bound death receptor molecule Fas resulted increased the rate of formation of proplatelet extensions and viable platelet release [50], hence supporting a role for extracellular death receptor ligand directed activation of initiator caspase. In contrast, however, a recent study indicated that ligation of Fas, instead of aiding platelet formation, triggers caspase-8 mediated cell death [53]. Another death receptor molecule, TRAIL, was also associated with megakaryocytic cell development. TRAIL induces a modest increase in the CD41 expressing cells in vitro in a population of CD34- derived megakaryocytes and an increase in the number of cells with multilobulated polyploidy nuclei [54].
Later it was demonstrated that throughout the process of megakaryopoiesis there is continuous expression of Bcl-xL [51]. Based on this, megakaryocyte differentiation was studied in Bcl-2 over-expressing transgenic mice model that are resistant to caspase activation. This study demonstrated a role for caspase activation in the early differentiation phase rather than at the proplatelet forming terminal differentiation phase. A parallel study was conducted on two other lineages of hematopoietic cells, erythroid and GM colony formation from c-kit+/Lin progenitor cells. As expected, the addition of z-VAD-fmk, a broad spectrum caspase inhibitor, significantly inhibited erythroid colony formation. Bcl-2-overexpressing MKs showed an ability to form proplatelets that was comparable to WT MKs. This study also showed that the addition of z-VAD-fmk had no effect on proplatelet production [53]. The apoptosis related factor Bim, a BH3 only domain member of Bcl-2 family, plays a role in cell cycle progression in megakaryopoiesis. Bim deficient progenitors are less capable of proliferating and differentiating into MKs in response to TPO [55].
Caspase-9 is the initiator caspase involved in mitochondria mediated apoptosis. The intrinsic apoptosis pathway activation is essential for platelet shedding [49], for which caspase-9 is required. However, conflicting results from a later study suggested that caspase-9 is not essential for the terminal differentiation of megakaryocyte; platelet production occurred normally in caspase-9 deficient fetal liver cells, indicating that the apoptotic caspase cascade is not mandatory for platelet production [56]. Although the authors ruled out the possibility of activation of the intrinsic pathway, they suggested a possible role for other caspases or other proteolytic enzymes in proplatelet formation by megakaryocyte. More recently a study was conducted on mice with deletion of caspase-8, BAK, and BAX in megakaryocytic cells. The authors found that platelet formation was uninterrupted and they also rescued mice megakaryocytes from apoptosis by introducing cytomegalovirus. The authors concluded that apoptosis, both intrinsic and extrinsic, is dispensable for platelet production [57].
Go to :
APOPTOSIS IN MONOCYTE TO MACROPHAGE DIFFERENTIATION
Macrophages are sentinels of the innate immune response. They are responsible for the identification and removal of apoptotic bodies, microbes, cancer cells, and other foreign particles, through a process known as phagocytosis, thereby inducing the production of inflammatory cytokines. Monocytes differentiate into a variety of macrophages, including osteoclasts in bone, microglia in central nervous system, Kupffer cells in the liver, and dendritic cells. Monocytes originate from a common myeloid progenitor. The inflammatory responses regulated by these cells are mediated by the transcription factor nuclear factor kappa-B (NF-κB). Recent studies have shown the role of NF-κB in the survival of monocytes. Embryonic macrophages from NF-κB null mice are susceptible to TNF-α induced cell death [58]. Macrophages are tolerant of apoptotic signals induced by Fas and TNF-α death receptor ligation as Fas associated death domain-like interleukin 1β converting enzyme (FLICE)-inhibitory protein (FLIP), a negative regulator of Fas mediated apoptosis, was up regulated during monocyte to macrophage differentiation [59]. Mcl-1, a Bcl-2 family member, essential for maintenance of mitochondrial membrane integrity in the absence of an external death inducing ligand, and regulate macrophage survival [60]. Rebe et al. demonstrated that caspase 8 activity is required for the differentiation of peripheral blood monocytes into macrophages [13]. Caspase-8 activation involves an adaptor molecule FADD, serine threonine kinase RIP1, and FLIP. FADD overexpression enhances macrophage differentiation; however inhibition of Fas mediated apoptosis does not inhibit differentiation [61].
Go to :
CONCLUSION
Apoptosis is a genetically controlled mechanism that plays a central role in maintaining cell number by dismantling unhealthy cells. Caspases are the major mediators of apoptotic cell death and play significant roles in in several non-apoptotic functions, including cell-cell communication, cytokine maturation, and inflammation. Apoptotic caspases are involved in cellular differentiation in several cell types. These caspases are transiently expressed in a regulated manner so that the activation does not result in cell death. In several cell types, caspase activation is mandatory for cellular differentiation. Caspase activation in the differentiation of erythrocyte is a well-established phenomenon, whereas, in megakaryopoiesis, both intrinsic and extrinsic pathways of apoptosis are dispensable. More in depth studies are needed to understand the role of caspase activation in the differentiation of these cell types.
Go to :
 XML Download
XML Download