

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Precursor B-cell acute lymphoblastic leukemia (B-cell ALL) is the most common neoplasm in children and is characterized by excessive proliferation of abnormal and undifferentiated B-cells. Chromosomal translocations that frequently involve genes encoding hematopoietic transcription factors (TFs), such as ets variant 6-runt-related transcription factor 1 (ETV6-RUNX1) t(12;21), mixed-lineage leukemia (MLL), and transcription factor 3-pre-B-cell leukemia homeobox 1 (TCF3-PBX1) t(1;19) translocations, are the hallmarks of B-cell ALL. Other genetic aberrations commonly detected in hematopoietic TF genes include point mutations, deletions and amplifications [1]. Genetic abnormalities in crucial regulators of B-lymphocyte differentiation are present in 40% cases of pediatric B-cell ALL [2], indicating that affected hematopoietic TFs are important for B-cell leukemogenesis.
Besides genetic aberrations, epigenetic mechanisms are involved in the development of leukemia [3]. The best-described epigenetic mechanism is the methylation of CpG islands in promoter regions that regulates gene expression. Several microarray-based studies have shown that pediatric patients with ALL show aberrant DNA methylation on a genome-wide scale, indicating a distinct DNA methylation pattern between patients with ALL and healthy individuals and among different B-cell ALL subtypes. Gene ontology analyses have shown that genes encoding TFs and other proteins involved in lymphocyte development are the most aberrantly methylated genes in pediatric patients with ALL [4, 5].
Previous studies on DNA methylation in pediatric patients with ALL have provided some insights on the promoter methylation status of genes encoding hematopoietic TFs. These studies have highlighted that early B-cell factor 1 and paired box 5 genes (EBF1 and PAX5, respectively), the master regulators of B-cell development, show no or negligible methylation [2], and that homeobox A and B cluster genes (HOXA and HOXB, respectively) show frequent methylation [6].
In this study we comprehensively analyzed promoter methylation and expression of selected hematopoietic TF genes homeobox A4 and A5 (HOXA4 and HOXA5), Meis homeobox 1 (MEIS1), T-cell acute lymphocytic leukemia 1 (TAL1), and interferon regulatory factors 4 and 8 (IRF4 and IRF8) in pediatric patients with B-cell ALL. TFs TAL1 (stem cell leukemia [SCL]), HOXA, and MEIS1 act in the early stages of hematopoiesis, i.e., during the formation of hematopoietic stem cells (HSCs) and early progenitor cells [7,8,9]. HOXA4 promotes the expansion of HSCs and regulates the initial stages of B-cell development [10]. IRF4 and IRF8 are critical factors that act in the later stages of normal lymphopoiesis and regulate B-cell proliferation and differentiation [11, 12].
MATERIALS AND METHODS
Patients
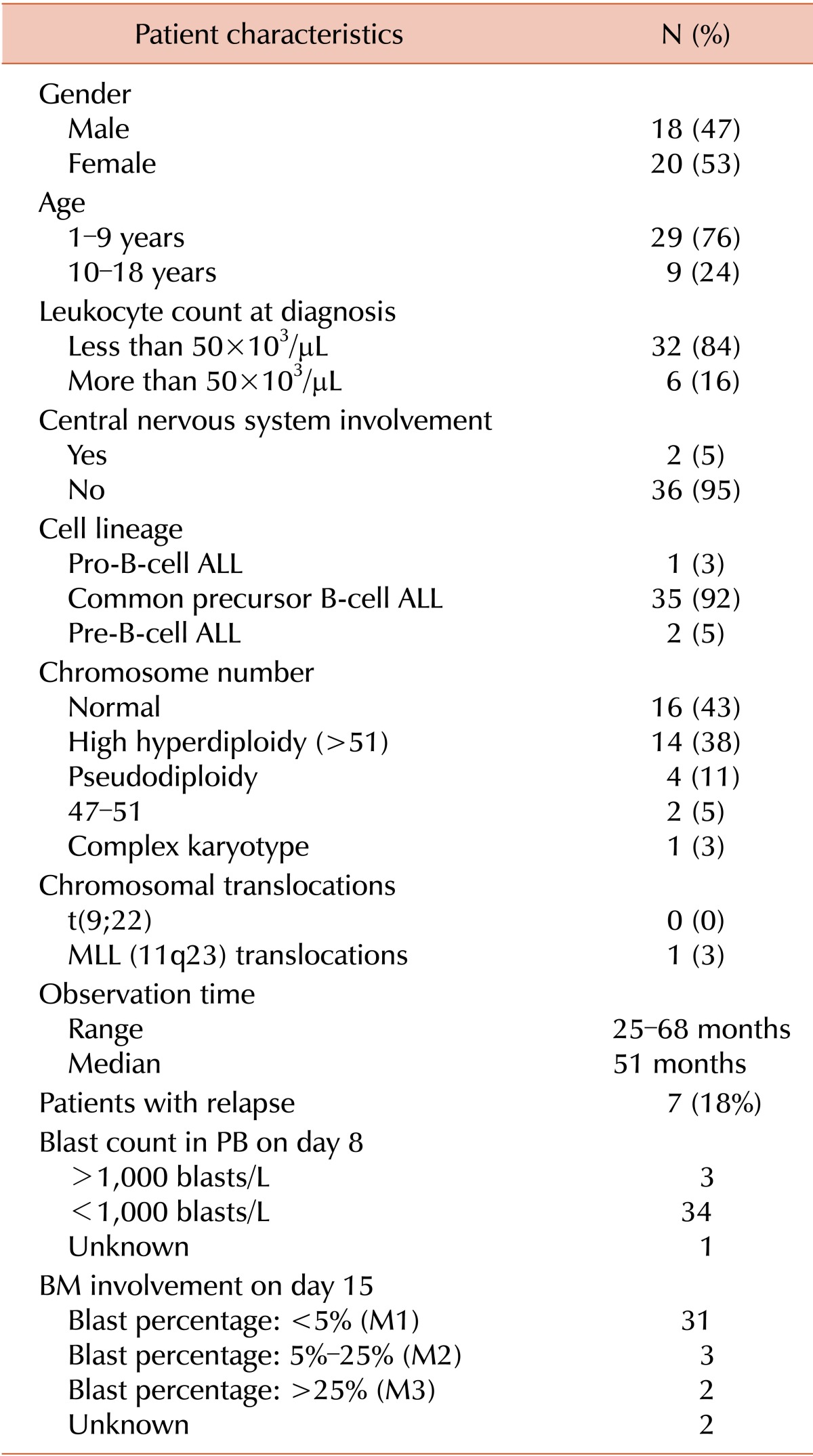
Blood samples were collected from 38 pediatric patients (age, 1-17 years) with ALL at the Medical University of Warsaw, Poland. The study was approved by the local ethics committee. Patient characteristics are listed in Table 1. Of the 38 patients, 35 had ALL B-common, 1 had ALL-proB, and 2 had ALL B-common/pre-B. Peripheral blood mono-nuclear cells (PBMCs) obtained from 20 anonymous healthy pediatric donors aged between 8 months and 17 years were used as controls.
Quantitative DNA methylation analysis
DNA was extracted from 27 Ficoll-isolated PBMCs and 11 whole blood samples using QIAamp DNA Mini Kit (Qiagen, Hilden, Germany), was quantified using NanoDrop (Thermo Scientific, Wilmington, DE, USA), and was treated with sodium bisulfite by using EpiTect Kit (Qiagen), according to the manufacturer's recommendations. Promoter methylation of IRF4, IRF8, HOXA4, HOXA5, MEIS1, and TAL1 was determined using quantitative methylation-specific polymerase chain reaction (qMSP) [13]. Briefly, methylation- specific primers and fluorescently labeled probes were designed using Methyl Primer Express Software (Applied Biosystems, Foster City, CA, USA; sequences listed in Additional file 1). ACTB region that does not contain CpG dinucleotides was used as a reference for normalizing the amount of DNA in each sample and was amplified using previously reported primers [14]. Polymerase chain reaction (PCR) was performed in 7.5 µL reaction mixture containing 1.5 µL bisulfite-converted DNA as the template, 2× SensiMix II Probe master mix (Bioline, Alexandria, New South Wales, Australia), 2.25 pmol of forward and reverse primers (4.5 pmol of both the primers for ACTB), and 1.9 pmol of fluorescent probes. Amplification was performed using 45 PCR cycles in 384-well plates by using 7900 HT Fast Real-Time PCR System (Applied Biosystems, Foster City, CA, USA).
Standard curves of known template concentration were used to quantify the PCR products. Samples were prepared using serial dilutions of plasmid DNA containing methylated inserts of the genes of interest. These recombinant plasmids were constructed by amplifying standard methylated genomic DNA (Qiagen) with primers for each selected gene and by cloning the PCR products with StrataClone Kit (Agilent Technologies, La Jolla, CA, USA). The recombinant plasmids were amplified in bacteria, purified using Plasmid Mini Kit (A&A Biotechnology, Gdynia, Poland), and quantified using Quant-iT PicoGreen (Invitrogen, Paisley, UK).
Percentage DNA methylation (percentage of methylated reference [PMR]) was calculated by dividing the gene of interest:ACTB ratio of sample by the gene of interest:ACTB ratio of the methylated DNA control (Qiagen) and by multiplying with 100.
Analysis of gene expression
Expression IRF4, IRF8, HOXA4, HOXA5, MEIS1 and TAL1 was analyzed in 27 samples from patients with ALL and 20 samples from healthy pediatric controls. RNA was isolated from PBMCs by using RNAqueous-Micro Kit (Ambion, Austin, TX, USA) and from whole blood samples by using TRI Reagent BD (Sigma Aldrich, St Louis, MO) and was digested using DNase (Qiagen), according to the manufacturer's instructions. RNA concentration was quantified using NanoDrop. Next, 500 ng of RNA from each sample was reverse transcribed using SuperScript II Reverse Transcriptase (Invitrogen), and real-time reverse transcription- PCR was performed using 7900HT Fast Real-Time PCR System. PCR was performed in 5 µL reaction mixture containing 1.5 µL 10× diluted cDNA template, 2× SensiMix II Probe master mix, and 5'-FAM and 3'-MGB TaqMan probes (Applied Biosystems) following cycling conditions: initial denaturation at 95℃ for 10 min, followed by 45 cycles of denaturation at 95℃ for 15 sec and amplification at 60℃ for 60 sec. Ubiquitin gene (UBC) was used as a reference. Gene expression was calculated using 2-ΔCt method, where ΔCt is the difference between Ct value of the gene of interest and the reference. For presenting the expression of different genes in a single graph, the results were normalized so that the highest value of gene expression corresponded to the arbitral value of 100 (Fig. 1B).
Statistical analysis
Gene expression and percentage DNA methylation (PMR value) were treated as continuous variables and were analyzed using 2-sided Mann-Whitney U-test and Spearman correlation. Results of qMSP were categorized into binary data for descriptive statistical analysis. Samples from patients with ALL and healthy controls were classified as hypermethylated if the result of qMSP exceeded a PMR value of 1 (1% of methylation), except for genes in samples from healthy controls having a methylation level of >1%. In such cases, mean +2 standard deviations (SDs) of PBMC results from healthy controls was used as the threshold for classifying samples from patients with ALL as hypermethylated.
To assess the possible prognostic value of promoter methylation and expression of the selected genes, we analyzed their relationship with clinical prognostic indicators such as age of patients, white blood cell (WBC) count at the time of diagnosis, blast count in the peripheral blood (PB) on day 8 or in the bone marrow (BM) on day 15, and relapse during follow-up. Data collection was terminated on September 10, 2014.
Significance threshold was set at α=0.05. All statistical analyses were performed using GraphPad Prism (La Jolla, CA, USA).
RESULTS
Analysis of promoter DNA methylation and gene expression in patients with B-cell ALL
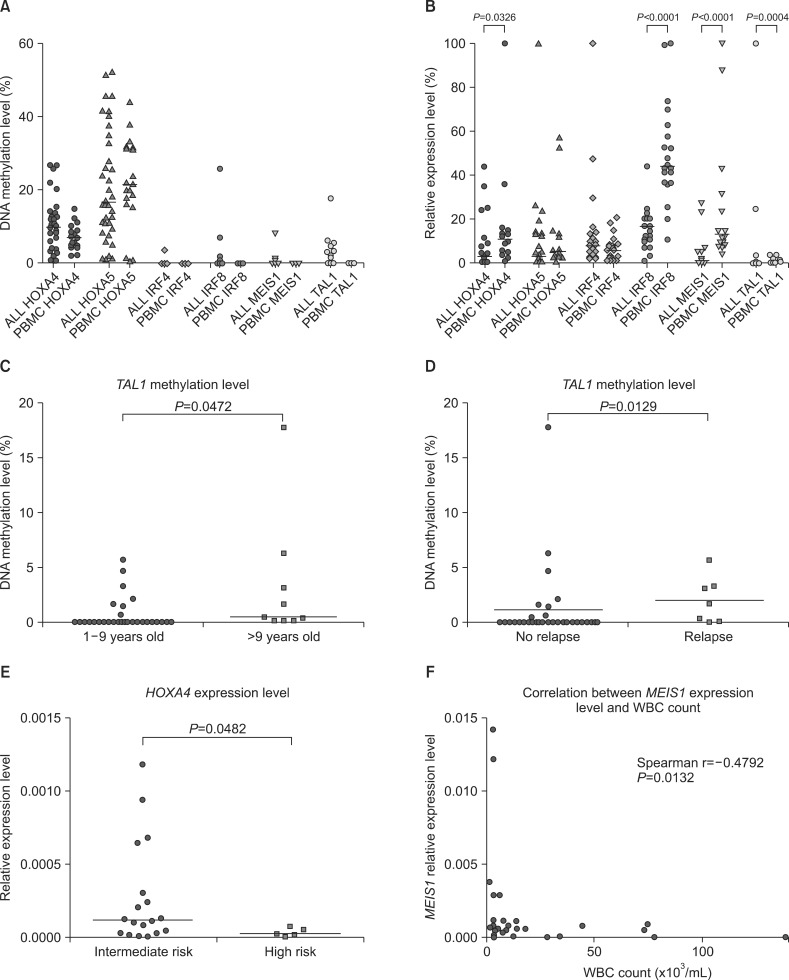
Promoter methylation level of HOXA4, HOXA5, MEIS1, TAL1, IRF4, and IRF8 was assessed in samples from 38 ALL patients and 20 healthy pediatric controls. TAL1, IRF4, IRF8, and MEIS1 were hypermethylated (PMR >1%) in 10 (26.3%), 1 (2.6%), 3 (7.9%) and 2 (5.3%) samples from patients with ALL, respectively, but not in samples from controls (PMR <1%; Fig. 1A). Promoter methylation level was very low in samples from patients with ALL, with only 1 sample showing >10% methylation of IRF8 and TAL1 and none of the samples showing > 10% methylation of IRF4 and MEIS1. Methylation of HOXA4 and HOXA5 was elevated in samples from controls (>1%). Therefore samples from ALL patients showing methylation level exceeding the defined threshold (mean +2 SDs of control samples) were classified as hypermethylated. HOXA4 and HOXA5 were hypermethylated in 6 (16%) and 2 (5%) samples from ALL patients, respectively (Fig. 1A). Importantly, notable differences were observed in the methylation level of both the HOXA genes in PBMCs from healthy controls. The observed PMR value ranged from 2.4 to 14.9 and from 1 to 44 for HOXA4 and HOXA5, respectively.
These 2 genes are located in the same chromosomal region. Interestingly, we observed an inverse correlation between promoter methylation levels of these genes in samples from ALL patients (Spearman correlation r=0.4415, P =0.0055; see Additional file 2).
Gene expression analysis showed significantly decreased expression of IRF8 (fold change of expression [FC]=-2.6, P< 0.0001), MEIS1 (FC=-6.4, P<0.0001), TAL1 (FC=-4.8, P=0.0004) and HOXA4 (FC=-2.1, P=0.0326) in samples from patients with ALL compared with that from healthy controls (Fig. 1B). Moreover, IRF4 expression was higher in samples from patients with ALL than in those from healthy controls; however, the difference was not statistically significant (Fig. 1B). Because DNA promoter methylation is generally associated with the regulation of gene expression, we evaluated the correlation between promoter methylation and mRNA expression. However, no statistically significant correlation was observed.
Comparison of expression level of the genes of interest showed correlation among genes encoding TAL1, homeobox proteins, and interferon regulatory factors. Inverse correlation was observed between IRF8 and HOXA4 expression (Spearman correlation r=-0.6756, P =0.0004; see Additional file 2). However, IRF4 expression was positively correlated with HOXA5 and MEIS1 expression levels (Spearman correlation r=0.4992, P =0.0111 and r=0.3946, P =0.0461, respectively; see Additional file 2). TAL1 expression level was positively correlated with HOXA5, MEIS1, and IRF4 expression (Spearman correlation r=0.7557, P <0.0001; r=0.5868, P =0.002; and r=0.6342, P =0.0005, respectively; see Additional file 2).
Correlation among promoter methylation, gene expression, and demographic and clinical characteristics of patients
We assessed the correlation among DNA methylation and expression levels of the analyzed genes and demographic and clinical features of pediatric patients with leukemia. According to the National Cancer Institute (NCI) risk classification, patients with ALL who are aged 1-9 years and who have a WBC count of <50,000/µL are classified as standard risk patients while those aged ≥10 years and who have a WBC count of >50,000/µL have a high-risk and show poor prognosis. In the present study, we observed slightly elevated TAL1 promoter methylation in patients aged ≥10 years (median value 0% vs. 0.6%, P =0.0472; Fig. 1C). MEIS1 expression was inversely correlated with WBC count (Spearman correlation r=-0.4792, P =0.0132; Fig. 1E). When patients were stratified according to the NCI risk classification, high-risk patients showed significantly decreased HOXA4 expression (FC=-4.16, P =0.0482; Fig. 1F).
No relationship was observed between promoter methylation and gene expression and blast count in PB on day 8 or blast count in BM on day 15. Interestingly, comparison of leukemic patients who experienced and did not experience relapse showed very slight but significant increase in TAL1 promoter methylation level in the samples of ALL with relapse (median value 0% vs. 1.693%, P =0.0129; Fig. 1D). No significant differences were observed in promoter methylation or gene expression with respect to gender and chromosome number.
DISCUSSION
In leukemic patients, genes encoding hematopoietic TFs frequently show both genetic and epigenetic aberrations [1, 2, 4]. Such abnormalities result in impaired hematopoietic differentiation and may contribute to leukemogenic transformation. In the present study, we evaluated promoter DNA methylation and expression levels of 6 genes encoding earlyand late-acting hematopoietic TFs in samples from pediatric patients with B-cell ALL and healthy controls.
HOXA cluster genes encode TFs that are important regulators of early hematopoiesis, predominantly at the stages of HSCs and progenitor cells [8, 10]. HOXA4 controls HSC and progenitor B-cell expansion, while HOXA5 controls specification of erythroid and myeloid lineages [10, 15].
Genome-wide analysis [4] and analysis of candidate genes [6, 16] showed that promoters of HOXA4 and HOXA5 are methylated in some patients with B-cell ALL. Frequency of patients with hypermethylated promoters of these hematopoietic genes was lower in our study than in a study by Strathdee et al. [6]. This may be because of the use of different analytical techniques for assessing DNA methylation (semiquantitative COBRA assay used by Strathdee et al.) and different thresholds for DNA hypermethylation. In our study, the threshold for hypermethylation was defined based on the methylation status of larger number of samples from healthy controls. HOXA4 and HOXA5 were methylated in all the samples from healthy controls, with variable methylation level, which in turn determined the high threshold value for classifying samples from patients with ALL. This variable and relatively high methylation of HOXA in healthy controls was previously observed in a study by Schotte et al., which focused on the relationship between miR-196b and HOXA expression [16].
We did not observe a direct correlation between promoter methylation level of HOXA genes and their mRNA expression level (both considered as continuous variables), as that observed in chronic lymphoblastic leukemia [17]. However, we observed increased promoter DNA methylation of HOXA4 and downregulation of its expression in some samples from patients with ALL compared with PBMCs from healthy controls. HOXA4 expression was significantly lower in high-risk patients than in intermediate-risk patients with ALL. This is in line with the previously reported prognostic role of the expression of HOXA family genes in respect of relapse-free survival [18]. However, no relationship was observed between relapse and HOXA4 expression in our study.
MEIS1 encodes a co-factor that enhances the DNA-binding ability of HOX proteins [19]. In patients with acute myeloid leukemia (AML) showing AML-ETO translocation and in T-ALL, MEIS1 promoter region was shown to undergo hypermethylation, resulting in the downregulation of MEIS1 expression [20, 21]. MEIS1 expression was lower in patients with B-cell ALL than in those from healthy controls. However, methylation of the MEIS1 promoter was only observed in 2 leukemic patients, suggesting that low MEIS1 expression was not because of promoter methylation.
Previous studies indicate that MEIS1 expression in leukemic patients is strictly associated with MLL mutation status. MEIS1 expression was significantly upregulated in a subset of pediatric ALL patients showing MLL rearrangements, but showed negligible expression in other patients with B-cell ALL [22]. Analysis of ALL-derived cell lines showed a relationship between MLL status and MEIS1 expression. None of the cell lines with wilt-type MLL expressed MEIS1; however, MEIS1 expression was observed in cell lines with MLL mutations [23]. These results are consistent with the results of our study. In our study, MEIS1 expression was significantly downregulated in a group consisting of only wild-type MLL patients. Although the biological effects of MEIS1 upregulation in leukemic patients with MLL rearrangements have been partially established, the impact of MEIS1 down-regulation on leukemogenesis in patients with B-cell ALL having wilt-type MLL remains to be elucidated.
Further, MEIS1 expression level was inversely correlated with WBC count. This can be interpreted as the expansion of leukemic clones which showed lower MEIS1 expression level than normal cells.
IRF4 and IRF8 play a key role in B-cell development, particularly during pre-B cell transition to immature B-cells, class-switch recombination and plasma cell differentiation [12, 24]. Epigenetic silencing of IRF4 and IRF8 is reported in AML and/or leukemic cell lines [25, 26]. However, no information is available on IRF4 and IRF8 promoter methylation and mRNA expression in pediatric ALL. In the present study, IRF8 expression was significantly lower in patients with ALL than in healthy controls. Probably this was not because of promoter methylation since only 3 patients (7.9%) showed increased IRF8 promoter methylation. This may be because of miRNA-mediated regulation of IRF8 expression, as that observed in AML [27].
Only 1 patient with ALL showed a slight increase in IRF4 promoter methylation; however, no statistically significant difference in gene expression between group of patients and controls was revealed. Adamaki et al. reported elevated IRF4 expression level in pediatric patients with acute leukemias including ALL B-common [28]. Our study comprised almost twice the number of patients with ALL B-common (N=35) included in the study by Adamaki et al. All the patients with ALL B-common, except 3 patients, showed IRF4 expression within the range observed in healthy controls, suggesting that IRF4 expression did not play a substantial role in the pathogenesis of pediatric ALL B-common.
TAL1 (SCL) is a hematopoietic TF predominantly expressed in HSCs, early progenitor cells, and erythroid and megakaryocytic precursor cells [7]. TAL1 promoter hypermethylation has been observed in patients with ALL as well as in ALL-derived cell lines [29]. In the present study, TAL1 promoter methylation was reported in 26.3% (10/38) patients with B-cell ALL but not in PBMCs from healthy controls. TAL1 expression was significantly downregulated in patients with B-cell ALL in contrast to that observed in patients with T-cell ALL in whom it is frequently overexpressed [30]. Patients aged ≥10 years (a criterion for NCI high-risk classification) showed slightly elevated TAL1 promoter methylation level. This increase in TAL1 promoter methylation was also observed in leukemic patients who experienced relapse during follow-up, suggesting its prognostic value. However, such a strong conclusion should be drawn with caution. The TAL1 promoter seems to be non-methylated in healthy controls; however, its methylation level in patients with ALL is low. Patients with ALL who experienced and did not experience relapse only showed 1.7% difference in the methylation of TAL1 promoter, which probably could not be detected using techniques that were less sensitive and accurate than real-time PCR, such as gel-based methylation-specific PCR. This implies low potential clinical and diagnostic value of TAL1 promoter methylation as a prognostic marker. Our result on TAL1 promoter methylation should be validated in further studies involving larger number of patients by using other quantitative DNA methylation assessment techniques such as pyrosequencing or MassARRAY spectrometry.
From the viewpoint of development-hematopoiesis, an interesting relationship between the expressions of different genes can be observed. Decrease in the expression of the late leukemogenesis related gene IRF8 in ALL may be because of the undifferentiated nature of the leukemic blasts. However, following this interpretation, we could expect undifferentiated leukemic blasts should show increased expression of early hematopoietic genes HOXA4, MEIS1 and TAL1, which is in contrast to that observed in the present study. However, our observations are in line with previously reported data. When the expression of HOX genes in patients with ALL was compared with that in different isolated immature B-cells populations, no clear relationship was observed between leukemic and potentially corresponding normal cells [18]. Leukemic cells showed distinct HOXA expression regardless of their stage of differentiation.
Irrespective of the dysregulation of leukemic blasts, some correlations were observed between the expressions levels of the studied genes that resembled their relationship in normal leukemogenesis. IRF8 expression was inversely correlated with HOXA4 expression level. HOXA4 is specifically involved in the early phases of hematopoiesis while IRF8 acts during B-cell maturation; thus, during normal B-cell differentiation, their activity is separated by time. Moreover, TAL1 expression was positively correlated with HOXA5 and MEIS1 expression levels in patients with ALL. This could be expected because these genes act concurrently in the early stages of hematopoiesis. However, it is intriguing that expression level of HOXA5, MEIS1, and TAL1 was positively correlated with that of another late-acting hematopoietic gene IRF4.
In conclusion, the proportion of pediatric patients with B-cell ALL included in this study showed aberrant promoter methylation in key hematopoietic genes. Methylation was predominantly observed for the promoters of HOXA4 and TAL1. Abnormal gene expression was also observed. However, no correlation was observed between promoter methylation and downregulation of gene expression. Methylation and expression level of some analyzed genes was correlated with demographic and clinical parameters such as age, WBC count, NCI risk classification, and relapse, which affect the prognosis of patients with ALL. These results suggested the potential prognostic role of TAL1 promoter methylation in pediatric B-cell ALL. Low frequency of aberrant promoter DNA methylation in IRF8, IRF4, MEIS1, and HOXA5 suggested a limited role of these genes in the pathogenesis of pediatric B-cell ALL and their low clinical significance.
 XML Download
XML Download