

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Cardiovascular diseases are major morbidity and mortality in the industrialized countries. In the United States, ischemic cardiovascular diseases are the number one cause of death [1]. Although numerous pharmacological and surgical therapies have been developed, many of the patients still exhibit grave symptoms and continue to show increasing heart failure or advance critical limb ischemia and the need for hospitalization [2-4]. Due to our limited therapeutic options, the demand for novel methods of treatment for ischemic cardiovascular diseases has been increasing. One of the major causes of ischemic cardiovascular diseases is atherosclerosis. This leads to the blocking of the blood vessels, which, in turn, needs a new vascular growth for the affected tissues to prevent or reverse disease progression. Endothelial cells are a major component of blood vessels and, thus, are essential for vascular growth; therefore, the treatment of ischemic cardiovascular diseases should primarily aim neovascularization through endothelial cells.
The notion of stem cells has attracted much attention due to their potential to develop new tissues. Although ethically controversial, embryonic stem cells (ESCs) have shown promising potential for tissue regeneration; therefore, ESCs have been extensively studied for cardiovascular regeneration in animal models for the past decade. On the contrary to their vast potential, ESCs trigger host immune response to allogeneic ESCs [5], have shown the risks of teratoma formation [6], and need to be ethically solved before clinical translation. Thus, induced pluripotent stem cells (iPSCs) have been discovered to circumvent such issues [7, 8]. iPSCs, however, still have many questions to answer regarding its benefits and impediments [9-13].
In addition, adult stem or progenitor cells have shown promising outlooks in treating cardiovascular diseases, especially bone marrow (BM)-derived cells. This discovery has led to a wider approach to cell therapy. BM-derived cells have shown regenerative effects in both preclinical studies and clinical trials. Because of their suggestive results, BM-derived cells have been widely investigated, particularly BM-derived mononuclear cells (MNCs), endothelial progenitor cells (EPCs), or mesenchymal stem cells (MSCs). BM-derived EPCs have displayed therapeutic potentials in myocardial and peripheral vascular ischemia in animal models [14-21] and positive results in a series of clinical trials using EPCs or similar BM cells to treat post-MI cardiac dysfunction and critical limb ischemia [22-34]. Contrarily, in other studies, BM-derived MNCs exhibited contradictory results in a similar subset of patients with myocardial ischemia [35-38]. As more clinical trials with MNCs, early EPCs, MSCs, and CD34+ cells are underway, more agreeable conclusion will be drawn in the near future.
Advances and expansions have been made involving therapeutic effects of BM-derived stem and progenitor cells. Formerly, transdifferentiation into vasculature and/or cardiomyocytes was thought to be the predominant mechanism responsible for cardiac regeneration or repair [17, 20, 21, 39-47]; however, recent studies exposed such notions were overestimated [48-50] and that humoral or paracrine effects are the main mechanisms. The humoral mechanism benefits regeneration or restoration of myocardium and peripheral vascular tissues by inducing neovascularization, protecting ongoing cell apoptosis and degeneration, and promoting regeneration of endothelial cells, cardiomyocytes, and smooth muscle cells through soluble factors or cell-to-cell contact [42, 51-58]. Numerous attempts to identify the soluble factors responsible for paracrine effects of MSCs led to discovering secreted frizzled related protein 2 (Sfrp2), an antagonist of Wnt signaling [59], which had a dose-dependent cytoprotective effect, with its concentrations up to 15 nM, in in vitro caspase activity assay. Additionally, in their follow-up study, the exogenous delivery of Sfrp2 to rat hearts, at therapeutic doses of 4 µg per heart, improved cardiac function in experimental MI [60].
Furthermore, the search of effector cells, using their markers that are not specific only to stem or progenitor cells, has led to the discovery of CD31+ cells, representing a subpopulation of BM- and peripheral blood (PB)-derived MNCs. These CD31+ cells were discovered to have higher angiogenic and vasculogenic activities and efficient neovascularization in hindlimb ischemia [61, 62]. Hence, based on this information, this review will cover the characteristics and possibilities of EPCs, BM-MNCs, and recently identified CD31+ cells.
Go to : 
ENDOTHELIAL PROGENITOR CELL (EPC)
Base on the idea that endothelial cells are major components in maintaining vascular homeostasis and in pathogenesis of a variety of diseases, endothelial cells have been explored and BM-derived circulating progenitor cells or angioblasts in human peripheral blood (hPB) have been discovered [63]. Because these cells had properties like endothelial cells (ECs) and could differentiate into ECs, BM-derived circulating progenitor cells were referred as EPCs. Based on the notion that BM-derived circulating progenitor cells contribute to blood vessel growth, Asahara and colleagues suggested the groundbreaking concept of postnatal vasculogenesis by showing that EPCs were incorporated into the vasculature in adult animals and formed new vessels in ischemic models [39]. In addition, the transplantation of EPCs was able to induce neovascularization and assist ischemic limb repair [14]. The concept of postnatal vasculogenesis has been widely accepted; however, due to the lack of specific markers and to diverse phenotypes, an accurate and precise identification of EPCs has not been confirmed, yet. In addition, the advance of technologies allowed the identifying of the role of EPCs in disease pathogenesis [64-67] besides a normal component of the formed elements of circulating blood [68].
1. Early EPC
Due to the lack of specific surface markers on EPCs, various BM cell fractions were short-term cultured in endothelial differentiation media to enrich EPCs. For example, CD133, which is displayed on immature hematopoietic stem cells (HSCs), was used for culture derivation of EPCs [69]. On the other hand, for therapeutic purposes, the total MNC population, which has been widely used to obtain EPCs, was short-term cultured for 4.7 days on vitronectin- or fibronectin-coated dishes and the attached, or adherent, cells were used as EPCs, though not all of the cells collected were considered EPCs [14, 39, 63, 70, 71]. These cells expressed endothelial-like characteristics by the uptake of acetylated low-density lipoproteins, the binding of lectins, the expression of several EC-specific proteins (VEGFR-2, Tie2, vascular endothelial [VE]-cadherin, von Willebrand factor, endothelial nitric oxide synthase [eNOS], and CD146), and a low proliferation rate. On the contrary, other studies have refuted the endothelial-like characteristics of EPCs because these cells also displayed monocyte/macrophage markers, such as CD45, CD11b, and CD11c [54, 72-74]. Recent studies have also, instead, referred these cells as angiogenic cells [74], for they contribute more to vessel formation through angiogenic effects rather than form ECs in vivo. EPCs are able to incorporate into the neovasculature and differentiate into ECs [20, 39, 63]. Also, Wang et al. [75] postulated that EPC containing cell population is inherently multipotent and may include smooth muscle progenitor cells. Additionally, other studies pointed that EPCs were able to differentiate into cardiomyocytes and smooth muscle cells in mouse [47] and rat [17, 21] models of MI; peripheral EPCs, mobilized by granulocyte-colony stimulating factor (G-CSF) administration and myocardial ischemia, have displayed cardiac-, muscle-, liver-, and neural-lineage markers [76, 77]. However, this evidence of extra-endothelial transdifferentiation is sporadic and not widely accepted after the original publications.
In one study, EPCs have been shown to transdifferentiate by fusing with cells of different lineage, such as newly formed cardiomyocytes in mice, administered with human EPCs, contained both human and mouse X-chromosomes [78]. However, only 3% of the ECs displayed any evidence of human-mouse cell fusion. In addition, most studies have shown little or no evidence of fusion between EPCs and cells of different lineages. In most recent studies, the transdifferentiation of EPCs has been shown to be possible but not prevalent.
Expression of certain factors and molecules, such as eNOS and inducible nitric oxide synthase (iNOS), which increase circulation by dilating capillaries [42], and pro-angiogenic and anti-apoptotic growth factors (i.e. hepatocyte growth factor [HGF], insulin-like growth factor 1 [IGF-1], and VEGF) [51], is one of the ways that EPCs contribute to tissue regeneration and preservation. The release of certain growth factors causes the recruitment of EPCs around the ischemic area [52] and, in turn, stimulates the cells to proliferate, migrate, and maintain the survival of EPCs and pre-existing mature endothelial cells [79].
In earlier studies, EPCs were often administered systematically; however, in recent studies, due to the low rate of retention, cells have been transplanted or injected directly onto the affected area. Transplantation of EPCs [14, 16, 20, 80, 81] facilitated neovascularization in animal models of acute myocardial ischemia, and analogous effects in humans [30, 82, 83].
2. Late EPC
Late EPCs, or endothelial colony-forming cells (ECFCs), were discovered from circulating MNCs that were collected after more than 2 weeks in conventional EPC culture. These cells may be cultured variably; however, they commonly have round morphology, fast cell proliferation rate, and express similar surface markers (EC makers) [73, 84]. ECFCs need to be studied much further and their culture systems developed more before any application to therapeutic or regenerative use, because they were able to neither convincingly show paracrine effects nor show vasculogenic effects without Matrigel™ plug assay [84, 85]. ECFCs' capability to differentiate into ECs and their regenerative and therapeutic effects on vasculature in ischemia need to be troubleshot. Furthermore, because there is no defined line between ECFCs and ECs, the possibilities that these cells are primitive circulating ECs or are ECs sloughed off from vessels still remain to be established.
Go to :
HEMATOPOIETIC STEM CELLS (HSCs)
HSCs are multipotent stem cells that have the capacity to give rise to all blood cell types, to commit to lymphoid lineages, and to self-renew [86]. In adult vertebrates, HSCs occupy < 1% of the total BM cells and constitute 1:10,000 cells in the myeloid tissue. Hematopoietic tissues contain cells in the long- and short-term regenerative capacities and committed multipotent, oligopotent, and unipotent progenitors. Currently, HSCs are one of the best characterized stem cells, due to their lack of markers indicating lineage commitment (lin-) and to the presence of combination of indicating markers, such as CD133, CD34, CD38, CD90 (Thy1), CD105, and CD117 (c-kit) [87-89].
HSCs have been applied for the treatment of hematologic diseases for over three decades. Furthermore, due to their multipotency, HSCs were applied for the regeneration of damaged myocardium [45, 46]. In these studies, HSCs were shown to be able to transdifferentiate into nonhematopoietic cells, such as cardiomyocytes, ECs, and vascular smooth muscle cells in infarcted heart [45, 46]. However, some studies have refuted that HSCs, represented by lin-negative, sca1-positive, and c-kit-positive, do not transdifferentiate into any cardiomyocytes, after transplantation, in a mouse MI model [48, 49]. While no other specific human HSCs have been used, human CD34+ [17, 27, 47, 90] and CD133+ cells [91-93] were used in experiments reporting that post-MI cardiac function has been improved, conversely, through non-cardiomyogenic effects. The need for the use of mobilizing cytokines, such as G-CSF, to collect sufficient amount of CD34+ or CD133+ cells expenses high cost for clinical application and may result in a risk to patients.
Go to :
BM MONONUCLEAR CELLS (BM-MNCs)
BM-MNCs are composed of a number of different types of cells, such as hematopoietic cells, fibroblasts, osteoblasts, myogenic cells, and endothelial-lineage cells [94]. BM-MNC transplantation facilitated neovascularization in animal models of acute myocardial ischemia [94, 95] and similar results in humans [96-99]. Because the monocyte/macrophage proportion of BM-MNCs expresses key angiogenic growth factors and cytokines (e.g., VEGF, basic fibroblast growth factor [FGF2], angiopoietin-1 [Ang-1], interleukin [IL]-1β, tumor necrosis factor-α [TNF-α]) and stimulates angiogenesis [94, 100, 101], this cocktail of angiogenic factors mixed in the population may cause paracrine effects that stimulate proliferation, homing, and differentiation of preexisting stem cells [102].
On the contrary to some of the beneficial effects exerted by BM-MNCs, lymphocytes and monocytes/macrophages express pro-inflammatory cytokines, such as IL-1, IL-2, IL-6, IL-12, interferon-γ (IFN-γ), lymphotoxin, and tumor necrosis factors (TNFs) [103-105], and may cause negative inotropic effect on myocardium. Also, because lymphocytes and monocytes/macrophages, which are inflammatory cells, are a large portion of the MNC population, myocardial damage after MI may be accelerated by these cells as well as severe hemorrhage or inflammation, instigated by the transplanted MNCs, could jeopardize the survival and transdifferentiation of transplanted MNCs [17].
Go to :
NON-TRANSDIFFERENTIATION THERAPEUTIC EFFECTS OF BM-DERIVED CELLS
The plasticity, true tissue generation, or transdifferentiation capacity of BM-derived cells has been widely debated, despite promising therapeutic potential [48-50]. Many recent studies suggest that the paracrine mechanism is the primary mechanism that accounts for therapeutic effects in cardiovascular diseases, not vasculogenesis or exogenous myogenesis because vasculogenesis and exogenous myogenesis rarely occur. BM cells release angiogenic factors VEGF, Chemokine (C-C motif) ligand 2, (CCL2, also known as MCP-1), FGF2, Angpt1, and Wnt [94, 106, 107]. It was reported that Wnt signaling mediate angiogenic effects of human fetal CD133+ cells on ischemic wounds [107] and that these growth factors stimulate angiogenesis, prevent tissue apoptosis or necrosis, and induce endogenous resident stem cell migration and proliferation via paracrine response [51, 55, 57, 58, 107, 108]. Recent studies reported that humoral effects can be attributed to both implanted cells and the target host tissues receiving the cell therapy [52, 108, 109]. Although fusion as a mechanism for transdifferentiation is well known [110, 111], unlike earlier studies [112, 113], recent studies have discovered that fusion is minimally involved in phenotypic changes of stem cells [58].
Go to :
HEMATOPOIETIC CD31+ CELLS
1. Role of CD31 in vascular biology
CD31, also known as platelet endothelial cell adhesion molecule (PECAM)-1, is a 130-kDa transmembrane protein consisting of six extracellular immunoglobulin folds. In its cytoplasmic domain, there are two immunoreceptor tyrosine-based inhibitory motifs (ITIM) for interactions with signaling molecules. CD31 is expressed on the cell surface of ECs and hematopoietic cells, such as monocytes, platelets, neutrophils, natural killer cells, megakaryocytes, and some T cells. CD31 mediates homotypic adhesion between adjacent ECs as well as between ECs and leukocytes [114, 115]. A role of CD31 in migration through ECs has also been reported for neutrophils and monocytes [116], and subsequently for numerous other cell types, including natural killer cells [117], hematopoietic progenitor cells (HPCs) [118] and certain subsets of lymphocytes [119, 120].
Cell-cell and cell-extracellular matrix interactions are two of the essential roles during blood vessel formation and regression. CD31 has been gaining attention as it is being recognized to suppress cell death through interactions, such as homophilic interactions of CD31 that are in between ECs and monocytes [121]. The interaction of CD31 in suppressing apoptosis suggests that CD31 releases pro-survival signals, and the interaction also has been reported to induce Akt, a serine/threonine protein kinase, phosphorylation. On the contrary to apoptosis suppression characteristics, CD31 also assist ECs in apoptosis through metalloproteinase-dependent cleavage and caspase-mediated cleavage of the cytoplasmic tail [122].
Additionally, given that it is expressed abundantly in ECs, CD31 has been shown to be involved in the initial stabilization and formation of cell-cell contacts at lateral junctions of ECs, the maintenance of a vascular permeability barrier, modulation of cell migration, transendothelial migration of monocytes and neutrophils, and formation of blood vessels in angiogenesis [114, 116, 123, 124]. Moreover, earlier studies have shown that CD31 formed a functional complex with cadherin 5, β-catenin, and F-actin to control EC tube formation [125]. However, a later study showed that CD31 was involved in the adhesion/signaling events necessary for the migration of ECs and subsequent tube formation during angiogenesis, independent of VE-cadherin [126].
2. Identification of specialized multimodal angio-vasculogenic cells
To advance cell therapy, it seems very crucial to target and identify cells that are specialized for humoral effects, since recent studies almost universally reported that the major therapeutic mechanism of BM-derived stem or progenitor cells is humoral or paracrine effect [52, 55-57]. Therefore, recent studies have used specific surface markers to isolate cells enriched in humoral activities from BM and/or PB [61, 62]. Using surface makers allows the cells to be isolated without cell culture, which saves costs, and safely evading any harmful effects that may come with using animal serum. In a series of studies, CD31 was used as a marker to find BM-derived cells that have multimodal effects, such as angiogenic, vasculogenic, higher adhesion, and almost exclusive hematopoietic stem/progenitor cell activities (Fig. 1).
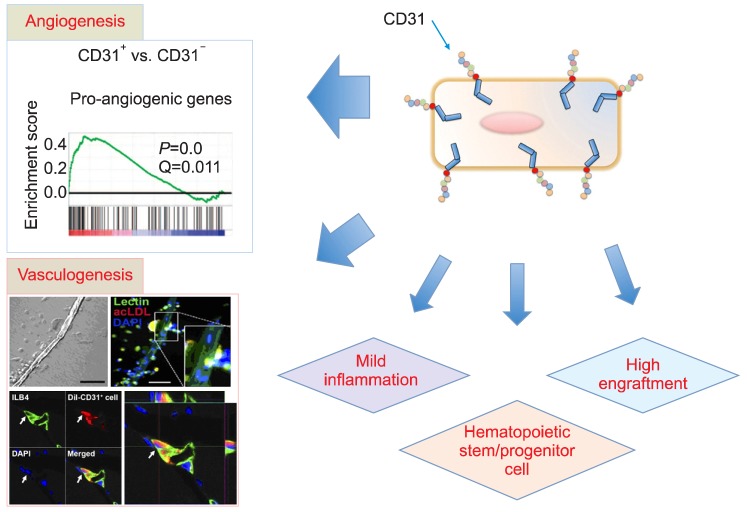 | Fig. 1The multimodal effects of BM-derived hematopoietic CD31+ cells. BM-derived CD31+ cells highly expressed pro-angiogenic genes (angiogenicity). BM-derived CD31+ cells showed a high number of EPCs and formed a tubular structure (vasculogenicity). The high adhesion capacity and low inflammatory activity of the cells allowed high engraftment and mild inflammation, respectively. The hematopoietic stem and progenitor cells were enriched in CD31+ cells (HSC and HPC activity) [61, 62].
|
3. Angiogenic activities
Based on genome-wide gene expression analyses of mouse BM (mBM), human BM (hBM), and hPB, angiogenic genes were enriched in CD31+ cells when compared with CD31- cells [61, 62]. Compared to their negative counterparts, hBM-CD31+ cells expressed higher levels of heparin-binding EGF-like growth factor (HBEGF), which is active in vascular smooth muscle cell recruitment [127] and IL-8, a macrophage-derived mediator of angiogenesis [128]; mBM-CD31+ cells expressed high levels of Angpt-1, an eminent angiogenic growth factor [129, 130], and GATA2, a transcription factor that proliferates angiogenesis when activated [131]. Similarly, hPB-CD31+ expresses extremely high levels of neuropilin-1 (NRP1), which is a necessity in vascular development and a mediator in VEGF-dependent angiogenesis [132], compared with the negative cells.
4. Vasculogenic properties
The recurring questions that need to be addressed are whether hematopoietic cells can give rise to ECs [46, 133, 134] and what is the true identity of those cells. A majority of studies, including ours, have demonstrated that BM-derived cells can generate ECs. On the other hand, several studies have refuted such notion [48, 50, 135] by using MI models to prove vasculogenic effects [48, 49]. However, those refutations are inappropriate since a MI model accepts minimal amount of engraftment from any cell, including BM-derived cells. Other studies supported our notion that the retention of ESC-derived cardiomyocytes or cardiac stem or progenitor cells was minimal in a MI model as well [136-138]. MI hinders engraftment and survival of any implanted cells, such as constant motion of the heart, inflammation in acute stages of infarction, insufficient oxygen and nutrients, and tight junctions between cardiomyocytes. One study reported that it is common for EC transdifferentiation to occur in transplanted heart samples [139]. In other independent studies, with the correct conditions, such as suitable BM-derived cells with permissive models, BM-derived cells can give rise to ECs [140, 141]. Based on the results of these studies, cell types and/or environment is very crucial to generating ECs from BM or PB cells.
In recent studies on the vasculogenic effects of CD31+ cells [61, 62], compared with CD31- cells, mBM-, hBM-, and hPB-CD31+ cells gave rise to a large number of EPCs in culture and expressed EC-specific markers, such as von Willebrand factor, VEGFR-2, VE-cadherin, and CD31. In addition, hPB-CD31+ cells formed aggregates and, subsequently, underwent tubular structural changes within the round cell cluster by day 10 followed by the formation of complete linear tubular structure that mimicked in vivo vasculogenesis. This linear tubular structure had stained positive for lectin and took up acetylated human low-density lipoprotein, indicating EC characteristics. In vivo vasculogenesis studies had also been investigated. By using a mouse model of hindlimb ischemia, vasculogenic activities of CD31+ cells had been tested [61, 62]. However, due to the divisive notion of transdifferentiation potential of BM cells [48, 50, 54, 134, 140, 141], the confirmation methods used were rigorous and definitive: confocal microscopy with 3D reconstruction of multiple images were used to clearly demonstrate that a fraction of CD31+ cells were colocalized with the ECs within the vascular structure even up to 8 weeks after; flow cytometric analysis of enzymatically digested hindlimb tissues showed that up to 4% of the ECs in the ischemic tissues were derived from transplanted mBM- or hPB-CD31+ cells; fluorescent in situ hybridization of the digested tissues further confirmed the contribution of hPB-CD31+ cells into ECs [62]. This clear adaptation of definitive methods were the first to prove transdifferentiation of hematopoietic cells. Clearly, this experiment had proved that directly injected CD31+ cells give rise to functional ECs in ischemic tissues.
5. Higher adhesion and engraftment potential
Low retention of injected cells is one of the major problems that hinder the therapeutic effects of cell therapy because a stable engraftment and a steady survival are needed for optimal cell therapy. Studies reported that less than 25% of engraftment of transplanted neonatal cardiomyocytes were present within 24 hrs of MI in animal models [142] and, similarly, only 5% of CD34+ injected cells remained after 1 hr in human patients with MI [143]. Additionally, a majority of the directly injected, cultured early EPCs in myocardium post-MI had disappeared within a week [52]. Cell adhesion is also another crucial factor that can increase survival. Cells that adhere to extracellular matrix have higher chance of survival since they are able to avoid anoikis, an apoptosis caused by the lack of adhesion to extracellular matrix. CD31 was first found as an adhesion molecule [115, 144] and had been shown to mediate cell-cell interaction via hemophilic interactions between CD31-expressing cells [145]. Genome-wide gene expression data showed high levels of genes relevant to adhesion, chemokine production and reception, and extracellular matrix in CD31+ cells [62]. Furthermore, cell adhesion assay showed that CD31+ cells have the capacity to adhere to more matrix proteins, such as fibronectin, collagen, laminin, and vitronectin [61, 62], than CD31- cells can. In vivo samples analyzed by the confocal microscopy and FACS analysis confirmed higher engraftment and survival of injected CD31+ cells compared to the CD31- cells or MNCs.
Although CXCR4/SDF-1 is a recognized signaling axis to mediate cell engraftment and migration, the engraftment of CD31+ cells do not depend on this axis because, when the levels of CXCR4 were measured, there was no difference in the expression between CD31+ and CD31- cells. On the other hand, CD31 molecules and other adhesion molecules, ICAM4 and integrin α, which were expressed more in CD31+ cells than CD31- cells, could have been involved in the higher engraftment of CD31+ cells. The higher engraftment of CD31+ cells would allow mBM- and hPB-CD31+ cells to increase angiogenic and vasculogenic capabilities.
6. Enriched HSCs & hematopoietic progenitor cells (HPCs)
According to the gene expression studies, hematopoietic stem and progenitor cell gene levels were reported to be higher in mBM-CD31+ cells and in hBM-CD31+ cells than in their corresponding negative cells, suggesting that more hematopoietic stem and progenitor cells are expected to be found in CD31+ cell population. Concurrent with this hypothesis, according to in vitro colony-forming assay and in vivo BM cell transplantation, hematopoietic stem and progenitor cells are almost exclusively found in mBM-CD31+ cells; also, more than 90% of HSCs, multipotent progenitor cells, common lymphoid progenitor cells, and common myeloid progenitor cells in mBM express CD31 [61]. Similarly, in humans, CD31 was expressed in 99.8% of CD34+CD133+ and 89% of CD34+CD133- hBM cells, and clonogenic HPCs are found in hPB-CD31+ cells [62]. These findings suggest that HSCs and HPCs are heavily enriched in the CD31+ cell population.
There is some possibility that HSCs or HPCs included in the CD31+ population could have played a role in ischemic tissue repair because CD34+ cells, which include most HSC and EPCs, were known to be effective for treating ischemic cardiovascular diseases in animal models and human patients. Since the therapeutic effects of hBM-CD34-CD31+ cells and hBM-CD34+ cells were reported to be similar in improving mouse limb ischemia, it is more likely that non-HSCs could play a significant role in improving limb ischemia and/or therapeutic neovascularization.
Go to :
CLINICAL APPLICATION
CD31 is the first marker that was used to isolate BM or PB cell subpopulation based on recent discovery on the mechanisms. With such isolation methods, cell mobilization can be avoided, and more effective, safe, and promising cell therapy can be developed to treat patients with advanced ischemic cardiovascular diseases. Approximately 30-35% of the total MNCs from hPB are CD31+ cells, which make them a better selection over either CD34+ or CD133+ cells [146, 147]. Due to such characteristics, the use of mobilizing agents, such as G-CSF, is not required for CD31+ cell collection [26, 146]. Moreover, unlike cultured EPCs or MSCs, CD31+ cells do not need cell culture, which can develop harmful side effects. The current cell culture condition of EPCs and MSCs use fetal bovine serum that may develop the risk of disease transmission through viral, prion, and zoonose contamination or immunological effects [148]. Similarly, the uncultured BM-derived MNCs or unfractionated cells are deemed inferior to CD31+ cells, because BM-derived MNCs include unnecessary cells that have potential risks, such as calcification [149] and aggravation of ischemia [150], while no such adverse effects have been reported in CD31+ cells [61, 62]. Therefore, after having isolated CD31+ cells from nonangiogenic and highly inflammatory CD31- cells, CD31+ population can be utilized for higher therapeutic uses with smaller risks.
Go to :
CONCLUSION
More investigation is needed to identify more angiogenic and/or vasculogenic cells for better efficiency and potency, since CD31+ cell population is heterogeneous, including T and B lymphocytes and myelomonocytic cells. Since CD31+ cells have been proven experimentally, the following studies should be clinical trials, starting with patients with critical limb ischemia or non-healing wounds. Also, since peripheral tissues provide more room for engraftment and less hostile environment for survival, critical limb ischemia should be an optimal introductory target to treat with CD31+ cells. Furthermore, with success, cell therapy with CD31+ cells could be expanded to treating other ischemic cardiovascular diseases, such as MI and stroke.
After a decade of investigating and experimenting different perspectives of cell therapy, the importance of humoral and paracrine mechanism, host environment in cell engraftment and therapeutic effects, and selected cells has been discovered and such knowledge has caused us to rethink and revamp our approach to cell therapy. If tissue generation from transplanted cells is not needed, instead of using stem or progenitor cells that are few and require much handling, using specialized angiogenic and/or vasculogenic effector cells and tailored treatment based on specific diseases should be the next aim in therapeutic effects.
Go to :
 XML Download
XML Download