

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Mesenchymal stem cells (MSCs) are known to have significant potential for cell therapies, including cell-based transplantations and regenerative medicine, owing to their homing ability, engraftment efficiency, and therapeutic value following a homing event, as evidenced by differentiation into tissue-specific cell types [1, 2] and potent immunosuppressive effects [3, 4]. However, the results have been variable in part, which may reflect the intrinsic heterogeneity of MSCs, as well as differences in culture conditions for isolated MSCs.
MSCs exhibit heterogeneous characteristics with regard to morphology, proliferation rate, and secreted factors. Previous studies have attempted to characterize the heterogeneity of these cells, and preclinical trials using cells based on the intrinsic heterogeneity have been conducted [5-9]. Numerous attempts have been made to develop more specific procedures for the isolation and preparation of appropriate cells from this heterogeneous cell population. In particular, ex vivo expansion of MSCs is used for developing and maintaining the cells used for cell therapy. The methods used to expand and characterize the cells are critical factors in preparing MSCs. However, adequate protocols for preparing and characterizing MSCs have not yet been standardized. Although ex vivo expansion of MSCs is one of the alternative strategies for overcoming intrinsic heterogeneity, several recently studies have shown that low initial plating densities could be beneficial for the optimal ex vivo expansion of MSCs and their subsequent differentiation [10-12].
Podocalyxin-like protein (PODXL), α6-integrin, and α4-integrin are known to contribute to hematopoietic stem cell homing to the bone marrow (BM) [13], as well as the clonogenicity of MSCs and their migration to damaged tissues [5]. Leukemia inhibitory factor (LIF) also regulates humoral and cellular immune responses in damaged tissues [14] and induces the in vivo expansion of BM progenitor cells that accelerates hematopoietic reconstitution [15]; thus, it supports hematopoiesis and enables the maintenance of a highly enriched, competitive, and repopulation of stem cells [16].
In this study, we explored differences in the expression of PODXL, α4-integrin, α6-integrin, and LIF genes that play a crucial role in cell migration and tissue regeneration in BM-MSCs expanded under various seeding densities and cultivation times. In addition, we confirmed the expression of stemness and immunomodulatory genes in BM-MSCs under appropriate culture conditions under which those genes were highly expressed. Ex vivo expansion of MSCs maintained for an adequate seeding density and culture time after plating at low cell density could provide a promising strategy for enhancing regenerative properties of MSCs.
MATERIALS AND METHODS
Isolation and culture of human BM-MSCs
The Institutional Review Board of Samsung Medical Center approved this study. After obtaining written informed consent from the patients, we obtained iliac crest BM aspirates from normal adult volunteers. Mononuclear cells were isolated from normal BM aspirates by using Ficoll-Hypaque (Histopaque-1077; Sigma-Aldrich, St. Louis, MO) density gradient centrifugation. Cells were plated at 3×105 cells/cm2 in low glucose Dulbecco's Modified Eagle's Medium (LG-DMEM; Invitrogen-Gibco, Rockville, MD) containing 10% fetal bovine serum (FBS; Invitrogen-Gibco) and 100 U/mL penicillin/streptomycin (Invitrogen-Gibco). After 24 h, non-adherent cells were removed. Adherent cells were cultured for an additional 5-10 days, until they were ~70% confluent.
Flow cytometry
Antibodies against the human antigens CD14, CD34, CD45, CD73, and CD90 were purchased from Becton Dickinson (Franklin Lakes, NJ). Antibodies against CD105 were purchased from Ancell (Bayport, MN). A total of 5×105 cells were resuspended in 0.2 mL PBS and incubated with fluorescein isothiocyanate- or phycoerythrin-conjugated antibodies for 20 min at room temperature. The fluorescence intensity of the cells was evaluated by flow cytometry (FACScan; Becton Dickinson), and data were analyzed with CELLQUEST software (Becton Dickinson).
Differentiation of MSCs
Osteogenic differentiation
First passage MSCs were plated at 5×105 cells/well in 6-well plates in LG-DMEM containing 10% FBS, allowed to adhere overnight, and replaced with LG-DMEM containing 10% FBS supplemented with 0.1 µM dexamethasone (Sigma-Aldrich), 10 mM β-glycerolphosphate (Sigma-Aldrich), and 100 µM ascorbate-2-phosphate (Sigma-Aldrich). The medium was changed every 3 days. After 14-21 days, osteoblast differentiation was determined by alkaline phosphatase expression.
Adipogenic differentiation
As described above, cells were cultured for 14-21 days after reaching confluence in LG-DMEM containing 10% FBS, 1 µM dexamethasone, 500 µM isobutyl methylxanthine (Sigma-Aldrich), 100 µM indomethacin (Sigma-Aldrich), and 10 µg/mL insulin (Sigma-Aldrich). Adipogenic differentiation was evaluated by detecting cellular accumulation of neutral lipid vacuoles via staining with Oil-red O (Sigma-Aldrich) solution.
Chondrogenic differentiation
A total of 1×106 cells were pelleted in a 15-mL tube by centrifugation at 300 × g for 5 min. Pelleted cells were cultured for 14-21 days after reaching confluence in LG-DMEM containing 1× insulin-transferrin-selenium (ITS; Invitrogen-Gibco), 1 mM sodium pyruvate (Invitrogen-Gibco), 0.1 µM dexamethasone, 397 µg/mL ascorbate-2-phosphate, and 10 ng/mL transforming growth factor-β1 (R&D Systems, Minneapolis, MN). Chondrogenic induction was evaluated at 80% confluence by detecting extracellular accumulation of chondrocyte matrix by toluidine blue (Sigma-Aldrich) staining.
Cell counting
Second-passage MSCs were used throughout these studies. MSCs seeded at a density of 200, 1,000, and 5,000 cells/cm2 were observed on day 5, 10, and 15 of culture with an inverted microscope (Olympus CK40; Olympus, Melville, NY). The total cell number of MSCs was determined using a hematocytometer (Marienfeld, Germany).
RNA extraction and RT-PCR analysis
Viable first-passage MSCs were plated at 200, 1,000, and 5,000 cells/cm2 and incubated for 5, 10, and 15 days each. To obtain interferon (IFN)-γ-treated MSCs, MSCs plated at 200 cells/cm2 that were cultured for 10 days were incubated in the presence of 200 IU/mL IFN-γ (R&D Systems) for 48 h. Total RNA was isolated from 106 cells by using the QIAGEN RNeasy Mini Kit (Qiagen, Valencia, CA), which was used to perform semi-quantitative reverse-transcription PCR assays with a commercial kit (PrimeScript 1st strand cDNA synthesis kit; TaKaRa Shuzo, Shiga, Japan). Band intensity was measured using NIH image/ImageJ and normalized against that of GAPDH mRNA. cDNAs were amplified using the primers shown in Table 1.
RESULTS
Characteristics of BM-derived MSCs cultured at various seeding densities for different times
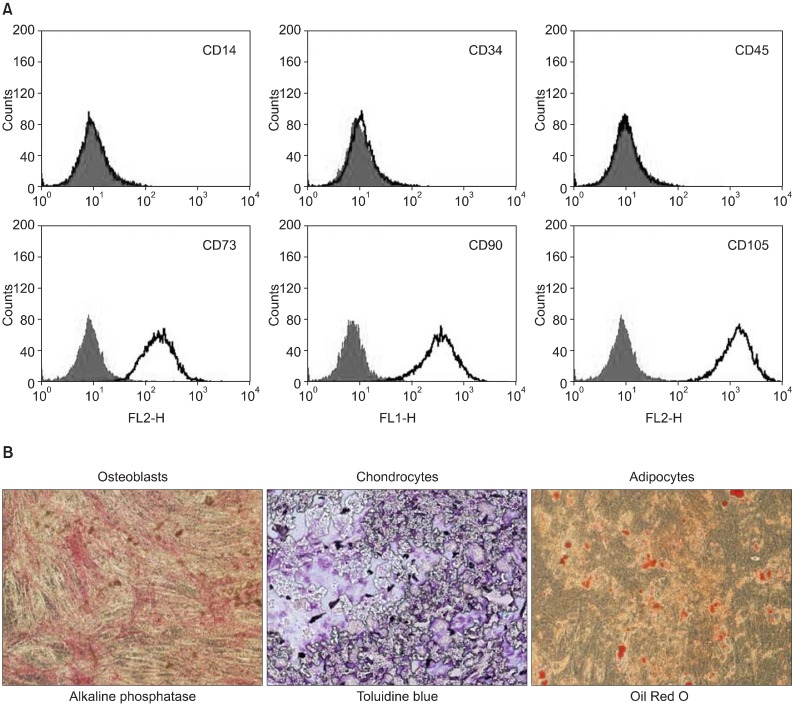
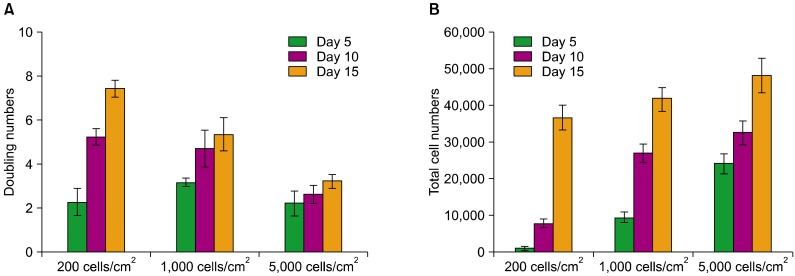
Human BM-derived MSCs were fibroblastic in shape. The cell surface antigen profiles of these cells after 2 passages in culture were analyzed by flow cytometry. The cells were positive for MSC-specific surface markers such as CD73, CD90, and CD105, but negative for CD14 and CD45 (Fig. 1A). The cells also exhibited mesenchymal differentiation potential, as assessed by culturing cells in osteogenic, chondrogenic, or adipogenic medium (Fig. 1B). Viable first-passage MSCs were plated at 200, 1,000, and 5,000 cells/cm2 and incubated for 5, 10, and 15 days (Fig. 2), and their yields were determined (Fig. 3). Cultures plated at a density of 200 cells/cm2 were ~50% confluent (7,800±1,200 cells/cm2) after 10 days, and most cells were thin and spindle-shaped. Cultures plated at 1,000 cells/cm2 were ~100% confluent (approximately 9,500±1,400 to 42,000±3,100 cells/cm2) after 5-15 days of culture. Cultures plated at 5,000 cells/cm2 were over-confluent, such that the cell yields were approximately 24,300±4,700 to 48,400±4,900 cells/cm2 after 515 days, and most cells showed extensive cell-to-cell contacts.
Comparison of gene expression in BM-MSC cultures relative to seeding density and culture time
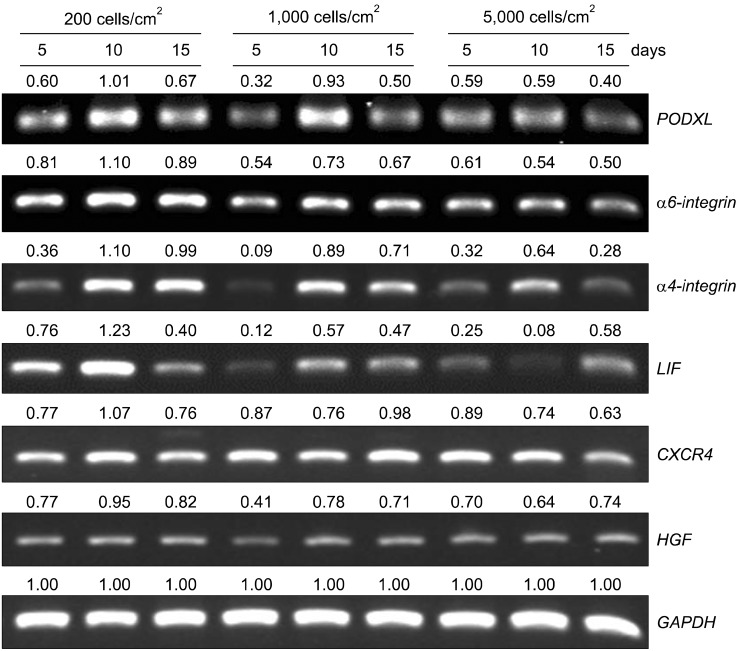
PODXL, α4-integrin, α6-integrin, and LIF are all linked to cell motility and migration [13, 17, 18]. After verifying the mRNA expression of PODXL, α4-integrin, α6-integrin, and C-X-C chemokine receptor type 4 (CXCR4) in MSCs plated at 500 cells/cm2, which is equivalent to a 50% confluent seeding density, the gene expression levels of MSCs plated at low or high densities were examined. RT-PCR revealed that PODXL, α4-integrin, α6-integrin, and LIF transcripts were present at the highest levels in 10-day MSC cultures seeded at a density of 200 cells/cm2 (Fig. 4). The PODXL mRNA levels were the highest in 10-day MSC cultures seeded at densities of 200 or 1,000 cells/cm2. The mRNA levels of α4-integrin, α6-integrin, and LIF were highest in cultures seeded at 200 cells/cm2, which were reduced when the cells were seeded at 1,000 or 5,000 cells/cm2. The mRNA levels of CXCR4 and hepatocyte growth factor (HGF) were not significantly different under any culture conditions.
Stemness and immunomodulatory properties of MSCs plated at 200 cells/cm2
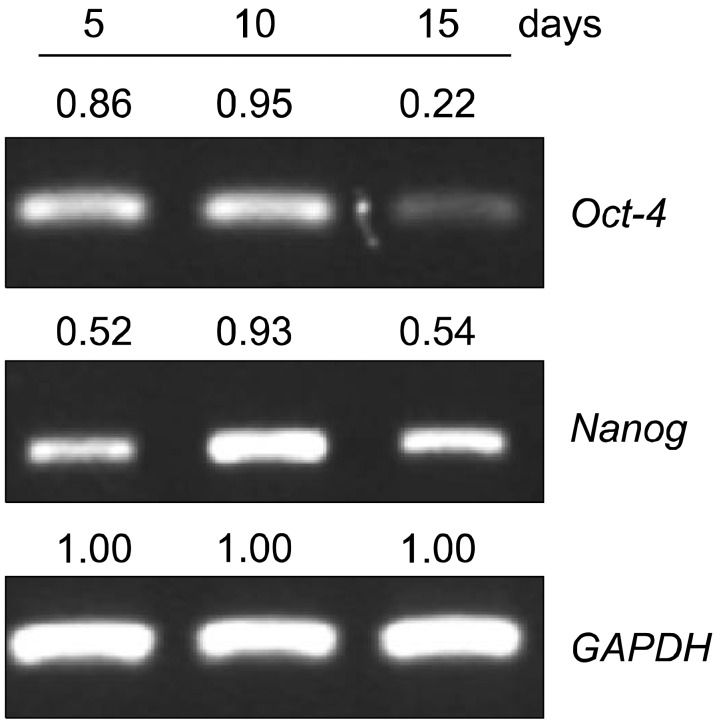
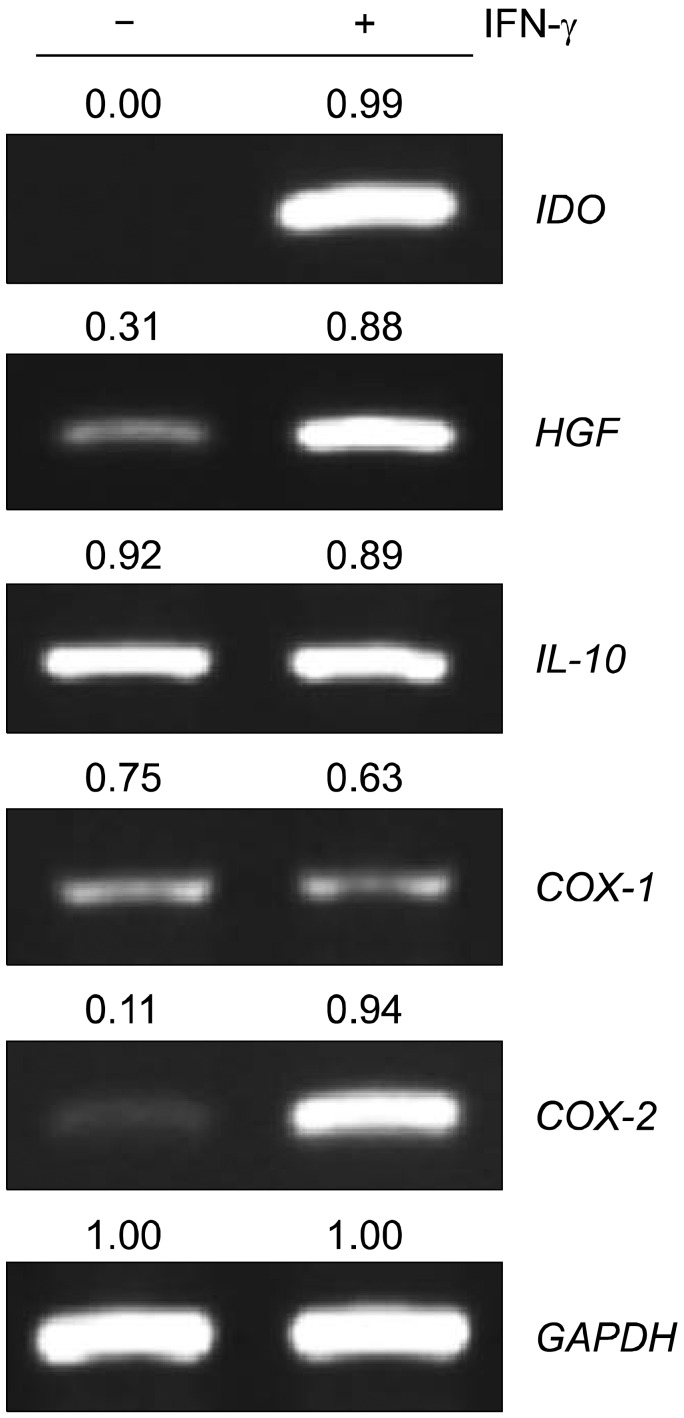
RT-PCR assays showed that Oct-4 and Nanog were expressed in MSCs seeded at a density of 200 cells/cm2 and cultured for 10 days (Fig. 5). To assess the immunomodulatory properties of these MSCs after 10 days in culture, the expression levels of immunomodulatory genes were analyzed. The mRNA expression levels of indoleamine 2,3-dioxygenase (IDO), cyclooxygenase (COX)-1, COX-2, and HGF was low in MSCs from all culture conditions, but that of interleukin (IL)-10 was high (Fig. 6). MSCs have been shown to retain their immunomodulatory properties in the presence of proinflammatory cytokines, for example, at inflammation sites in vivo that are rich in proinflammatory cytokines, especially IFN-γ [19, 20]. MSCs cultured for 10 days after plating at 200 cells/cm2 were incubated in the presence of 200 IU/mL IFN-γ for 48 h. IFN-γ treatment significantly upregulated IDO, COX-2, and HGF expression in MSCs seeded at 200 cells/cm2 and cultured for 10 days (Fig. 6). These data support the retention of an immunomodulatory role for MSCs.
DISCUSSION
The heterogeneity of MSCs and differences in conditions used to culture MSCs may be the main causes of the contradictory results from numerous studies and clinical trials. As a result, the methods by which the cells are expanded in culture and characterized are critical factors in terms of preparing MSCs with the desired features. By expanding MSCs ex vivo for 10 days after seeding at 200 cells/cm2, we obtained cells expressing PODXL, α6-integrin, α4-integrin, and LIF. Several studies have demonstrated that the "stemness" of MSCs, as characterized by their high proliferative potential and self-renewal capacity, is important for biomedical applications [7, 9, 21]. MSCs in low-density cultures contain a subpopulation of rapidly self-replicating cells [6-9] that could have a greater capacity to generate single-cell-derived clones through rapid proliferation compared to MSCs in high-density cultures. The surface protein characteristics of MSCs, including PODXL and α6-integrin, are known to reappear when subconfluent cultures are replated at low densities [5]; this finding indicates that surface proteins could be used to identify primitive cells in cultures. The genes for PODXL, α6-integrin, α4-integrin, and LIF were expressed at high levels in MSCs cultured at low density, but this does not imply that all MSCs plated at a low density will express these factors regardless of the culture time. Interestingly, the mRNA levels of these factors gradually increased over 10 days and then decreased by 15 days in culture. The expression level gradually changed with increasing seeding cell density. Therefore, the ex vivo expansion of MSCs plated at low densities must be maintained for an adequate culture time (e.g., 10 days).
MSCs expanded ex vivo for 10 days after seeding at 200 cells/cm2 showed higher gene expression of PODXL, α6-integrin, α4-integrin, and LIF. The expression of these genes could contribute to enhancing migration and intercellular communication, as well as homing of hematopoietic stem and progenitor cells to the BM and MSCs to damaged tissues [5, 13]. Confluent cultures of MSCs are useful for therapeutic preparations in terms of obtaining numerous cells, but these cells are more prone to produce lethal emboli when transplanted, possibly because they express cell adhesion proteins such as vascular cell adhesion molecule, and lack anti-cell adhesion proteins such as PODXL, and they therefore aggregate in the larger lung blood vessels [5]. Thus, the expression of these genes in MSCs may have a positive effect on both MSCs and their target cells in cell therapy settings.
Damaged tissues require the stemness and potent immunomodulatory properties of MSCs as a component of the therapeutic action of MSCs following a homing event for tissue reconstitution. The observed expression of Oct-4 and Nanog in 10-day cultures after seeding at a density of 200 cells/cm2 shows stem cell maintenance in the adult organism that is essential for tissue homeostasis and for initiation of native tissue regeneration and responses to injury [22]. Therefore, the identification of regulatory factors that maintain the "stemness" properties of MSCs is important for the practical purpose of producing sufficient quantities of MSCs for therapeutic applications. The anti-inflammation and immunomodulation exhibited by MSCs in their initiation of native tissue regeneration may also be important for tissue reconstitution in therapeutic settings. The inhibitory effect of MSCs on human mononuclear cell proliferation [20] and immunomodulatory gene expression of cultured MSCs when seeded at a low density indicates that MSCs retain their immunomodulatory properties in an inflammatory environment (i.e., in the presence of IFN-γ); this increases the possibility of protecting target stem cells and tissues, thereby supporting a possible use for MSCs in regenerative therapy under inflammatory conditions.
In conclusion, our results showed differences in the gene expression patterns of MSCs according to the seeding density and culture time. MSCs from 10-day cultures after seeding at a low density were efficiently expanded, expressed PODXL, α6-integrin, α4-integrin, and LIF, and retained their stemness and immunomodulatory gene expression. These data indicate that the seeding density and culture time are key factors in preparing MSCs for use in regenerative medicine. This study provides promising information on culture conditions appropriate for obtaining MSCs for cell therapy.

 XML Download
XML Download