

 PDF
PDF ePub
ePub Citation
Citation Print
Print


I. Introduction
Nowadays, bone tissue engineering is emerging as one of the important therapeutical approaches throughout all fields of regenerative medicine particularly the oral and maxillofacial area. In particular, various adult mesenchymal stem cells are being actively studied as potentially strong material of tissue engineering because it regenerates most similarly to natural osseous tissue1. Stem cell can self-renew by differentiating repeatedly; it has multi-lineage differentiation capability with which it can morph into a cell with a certain function depending on the environment. According to the environments, mesen chymal stem cells have the capability to differentiate into at least of three specific cell lines: osteo/odontogenic, adipogenic, and neurogenic cells2. These adult mesenchymal stem cells originate in various tissues such as bone marrow (bone marrow-derived stem cells, BMSCs) and adipose tissue (adipose tissue-derived stem cells, ADSCs); cells that originate with teeth and related tissues include the following: dental pulp stem cells (DPSCs), periodontal ligament stem cells (PDLSCs), stem cells from the apical papilla (SCAP), stem cells from human exfoliated deciduous teeth (SHED), dental follicle precursor cells (DFPCs), and dental papilla cells (DPCs)2,3. There are a lot of in vivo/in vitro experimental research studies reporting successful mineralization of mesenchymal stem cells that have been isolated and cultured for bone tissue regeneration4. To use mesenchymal stem cells for proper tissue regeneration, which is needed for pre-clinical/clinical experiments, however, the handling method of mesenchymal stem cells should be optimized. In this stage, the introduction of specific genes related to osteogenic/odontogenic differentiation can be an additional strategy for maximizing the effectiveness of stem cell use5. Therefore, in this study, we look into cases of maximization of differentiation capability by introducing specific genes related to osteogenic/odontogenic differentiation in recent literature; based on this, we discuss the direction of stem cell therapy using gene introduction in the oral and maxillofacial area.
II. Modulation of Osteoblastic/Odontoblastic Differentiation of Mesenchymal Stem Cells through Specific Gene Introduction
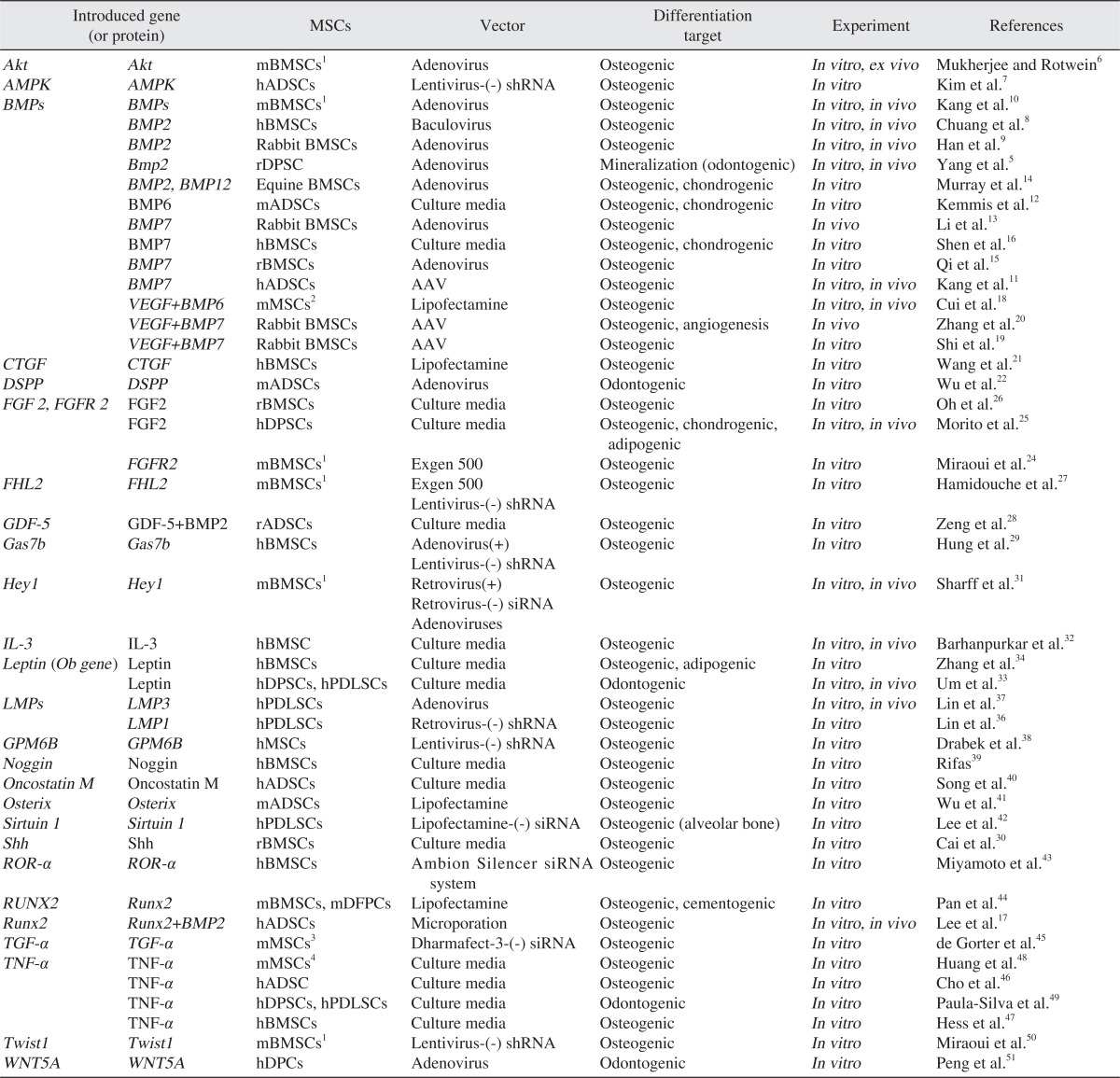
In this study, we considered literature published for the last five years and which reported the osteoblastic/odontoblastic differentiation capability of mesenchymal stem cells through specific gene introduction. As the genes introduced directly to stem cells in gene form or added in the culture medium of in vitro experiment in recombinant protein form or their protein product, the following were researched: Akt6, adenosine monophosphate kinase (AMPK)7, bone morphogenetic protein (BMP)5,8-16, BMP2 and Runt-related transcription factor 2 (RUNX2)17, BMP6/7 and vascular endothelial growth factor 165 (VEGF165)18-20, connective tissue growth factor (CTGF)21, dentin sialophosphoprotein (DSPP)22, fibroblast growth factor 2 (FGF2) or fibroblast growth factor receptor 2 (FGFR2)23-26, FHL2 (member of the LIM-only subclass of the LIM protein superfamily)27, growth and differentiation factor 5 (GDF5)28, growth arrest-specific gene 7b (GAS7b)29, sonic hedgehog (Shh)30, Hey131, interleukin-3 (IL-3)32, leptin33,34, LIM domain mineralization proteins (LMPs)35-37, neuronal membrane glycoprotein gene (GPM6B)38, noggin39, oncostatin M (OSM)40, osterix (Osx)41, sirtuin 1 (SIRT1)42, retinoic acid-related orphan receptor-α (ROR-α)43, Runx244, transforming growth factor β (Tgf-β)45, tumor necrosis factor-α (TNF-α)46-49, Twist150, and wingless-type MMTV integration site family (WNT)5a51 (Table 1). To express these genes, the following mesenchymal cells were used: BMSCc derived from mouse, rat, rabbit, horse, and human; ADSCs derived from human and mouse, and; DPSCs, PDLSCs, DFPCs, and DPCs derived from human and mouse. As the tool used to introduce the gene to cells, viral vector such as adenovirus, baculovirus, adeno-associated virus, retrovirus, lentivirus, and nonviral vector such as Lipofectamine 2000 (Invitrogen, Carlsbad, CA, USA), Exogen 500 (Euromedex, Mundolsheim, France), and Dharmafect-3 (Dharmacon, Lafayette, CO, USA) were used; over-expression through the introduction of the genes or, on the contrary, the inhibition of osteogenic/odontogenic differentiation by inhibiting the function of genes through the introduction of siRNA or shRNA, was reported. Moreover, there were research studies on modulation of differentiation by putting the specific genes in culture medium in the form of recombinant protein5-51. In the following, we wanted to explain focusing on the genes that were studied and reported a lot of times.
1. BMP
BMP plays an important role in stem cells' activity, adjusting the proliferation and differentiation of cells. Likewise, continuous studies show that BMP plays an important role in osteoblastic differentiation and bone formation. Belonging to the TGF-β superfamily, BMP consists of over 15 kinds in humans, and a few of them were reported as strong derivative for bone formation10. Kang et al.10 introduced 14 different kinds of BMPs to the C3H10T1/2 cell line, and discovered that the activity of alkaline phosphatase (ALP) had been improved when BMP2, 4, 6, 7, and 9 was overexpressed in C3H10T1/2 cell line - which is murine BMSC - using adenovirus. They also reported that ossified tissue masses had been formed unlike the negative control 5 weeks after the hypodermic injection of BMSCs to which BMP2 and BMP9 were introduced in an in-vivo experiment using nude mouse.
Chuang et al.8 stated that ALP activity and modulation of ossification in Alizarian red staining had been promoted as a result of the early gene introduction of BMP2 in human BMSCs using baculovirus and one more gene introduction six days after the first (supertransduction). According to them, when human BMSCs - to which genes were introduced - were put in alginate, and then injected into the belly area of nude mouse with CaCl2, they were observed to have started effectively differentiating into osteoblast two weeks after the injection; mineralized tissue and partial bone formation were also noted through eosin-hematoxylin dyeing and immunohistochemical staining 6 weeks after the injection. A similar result was found in the research on the gene introduction of BMP2 to DPSCs. According to Yang et al.,5 as a result of putting the BMP2 gene-introduced DPSCs in ceramic scaffold and implanting it into a nude mouse, they could observe high ossification effect, suggesting that 33±7.3% of the gap of the implanted scaffold had been filled with mineralized tissue 12 weeks later. A significant result obtained in this study is that the mineralized tissue cells can be seen as mineralized tissue similar to dentinum based on the high amount of DSPP and dentin matrix protein 1 (DMP-1) - which are indicators of dentinum - although they cannot be concluded as odontoblast5. Likewise, Li et al.13 studied the animal model whose mandibular damage had been repaired using gene-introduced stem cells. After forming a critical size defect in the mandible of a rabbit, it was divided into three groups - A, B, and C - for observation. For group A, BMP7-introduced rabbit BMSCs were implanted onto the nano-hydroxyapatite/polyamide (nHA/PA) scaffold; for group B, only BMSCs were implanted onto the scaffold. For group C as the control group, only scaffold was implanted. As a result of comparing bone tissue formation after 4, 8, and 16 weeks, group A showed excellent bone tissue formation in the 4th week and 8th week. There were no significant differences among those three groups in the 16th week, however. The result above proves that the gene introduction of BMP7 was effective in promoting bone formation in early treatment13.
In many ongoing research studies, other genes are introduced with BMP for the modulation of osteoblastic differentiation for effective bone regeneration. Specifically, some reports showed that the modulation of osteoblastic differentiation through the gene introduction of both BMP6/7 and VEGF as angiogenesis is also needed for bone regeneration. According to Cui et al.18 the bone-marrow-derived murine precursor cell line D1, which has multi-lineage differentiation capability through the gene introduction of both VEGF and BMP6, showed more increase of ALP activity and gene expressions of osteogenic differentiation than the cells wherein only BMP6 had been introduced. Likewise, in the in-vivo experiment wherein cells were implanted in polylactic-co-glycolic acid (PLAGA) scaffold and subsequently implanted into the belly area of a mouse, they reported that the experimental group - wherein the both genes were introduced - showed more bone formation and more angiogenesis than the group wherein just one gene was introduced or only D1 cell was implanted without gene introduction.
2. RUNX2
As the downstream target of BMP signaling and TGF-β1 signaling, RUNX2 is a major transcription factor that is essential in osteological development and modulation of differentiation17,44,52. Through the structural-functional analysis, it was reported that C-terminal with 5 amino acids (VWRPY) of RUNX2 suppressed the transcriptional activity of RUNX244. Pan et al.44 performed the gene introductions of full-length runx2 and mutant runx2 (runx2[M]) without VWRPY to murine DFPCs using Lipofectamine 2000. As a result of the experiment, the two experimental groups showed high expressions of genes related to the differentiation of osteoblast and cementoblast compared to the negative control group. In particular, the runx2(M) group showed high expression of osteocalcin (OC), osteopontin (OPN), collagen I (Col I), and cementum protein 23 (CP23) compared to the runx2 group. In mineralization assay, the two experimental groups wherein gene introduction was performed showed broader calcified area than the negative control group. In particular, the Runx2(M) group had more calcified area than the runx2 group44. Since Runx2 is connected closely with BMP signaling, Runx2 over-expression in murine BMSCs increased osteogenic differentiation during BMP-modulated osteoblastic differentiation, and the Runx2 knockdown reduced the modulated bone formation10. Lee et al.17 carried out the introduction of BMP2 and RUNX2 gene to human ADSCs using one bicistronic vector. In the analysis of calcium deposit conducted 14 days after the start of cell culture, the BMP2-introduced experimental group showed over three times' deposition amount than the negative control group; the BMP2-RUNX2-introduced group had somewhat less than three times' deposition amount than the BMP2-introduced group. As a result of 6 weeks' observation after the implantation of ADSCs implanted in scaffold into the belly area of a nude mouse, the negative control group wherein only ADSCs had been implanted showed no bone formation, whereas the BMP2-introduced group had immature bone. The BMP2-RUNX2-introduced group showed a developed angiogenesis as well as more matured bone. The results of the experiments above suggest that RUNX2 promotes bone formation in single gene introduction. Furthermore, it induces osteogenic differentiation and formation more effectively when introduced together with BMP210,17.
3. FGF2 and FGFR2
FGFs serve as a growth factor that plays an important role in the proliferation, differentiation, and survival of many tissue cells24. In particular, FGF2 (basic fibroblast growth factor) was proven to promote the healing of acute and chronic injury clinically and differentiate BMSCs into osteoblast24,25. The activity of these FGFs is closely linked with the FGFR. The combination of FGF and FGFR activates various signaling systems including the proliferation and differentiation of osteoblast by reducing the dimerization of receptor and phosphorylation of intrinsic tyrosine residues24. Oh et al.26 made collagen hydrogel by culturing the mixture of medium that included recombinant FGF2 and collagen solution containing BMSCs and observed FGF2 flowing out slowly over 30 days. As a result of the experiment, in BMSCs cultured in the collagen hydrogel matrix that included FGF2 and ALP, mineralization and expression of Col I, OPN, BSP, and OCN were more improved than the negative control group collagen hydrogel matrix without FGF2. Miraoui et al.24 gene-introduced the genetically modified and activated (MT) FGFR2 - which plays the role of promoting osteoblast in wild-type FGFR2 and Apert syndrome - to murine BMSCs. The result of this experiment showed high cell proliferation in the experimental group - wherein FGFR2 was introduced - particularly higher cell proliferation in the experimental group wherein MT FGFR2 was introduced. Moreover, in the experiment on the expressions of genes related to the differentiation into osteoblast and activity of ALP, the experimental group that had gene introduction showed higher results; in particular, the experimental group wherein MT FGFR2 had been introduced showed the highest differentiation into osteoblast24. Human DPSCs cultured in the medium containing recombinant FGF2 showed about eight times' expression of STRO-1, the stem cell marker, than the DPSCs cultured in the medium without FGF2. The experiment suggests that FGF2 promotes cell proliferation rather than cell differentiation and maintains cell's multi-lineage differentiation capability25. As a result of the transplant of DPSCs into scaffold and implantation in the body of mouse, however, calcified tissues were formed. Therefore, the research stressed that we cannot exclude the possibility of FGF2 inducing the differentiation of DPSCs into odontoblast25. According to Cucchiarini et al.23 the gene introduction of FGF-2 into human BMSCs using adeno-associated virus vector (AAV) had reduced the expression of gene-related osteogenic differentiation and mineralized tissues and had promoted the differentiation into cartilage. Regarding the influence of FGF2 on the differentiation of mesenchymal stem cells, careful planning of future research, i.e., minimizing unknown culture conditions, seems necessary.
4. TNF-α
TNF-α is a cytokine with various functions such as immune reaction, cell differentiation, proliferation and extinction, and adjustment of osteoporosis pathology. Moreover, as the factor in charge of catabolism, it promotes the activity of osteoclast but obstructs the activity of osteoblast46-49. The effect of TNF-α on mesenchymal stem cells is not clearly known, however. Whereas Li et al.53 reported that TNF-α suppressed the osteogenic differentiation of mesenchymal stem cells by preventing the expression of transcriptional coactivator with PDZ-binding motif (TAZ), Hess et al.47 claimed that the osteo genic differentiation of human BMSCs was promoted by vitalizing the signaling system of nuclear factor-kappa B (NF-κB). Huang et al.48 experimented on the ALP activity of the murine MSC ST2 cell, which was cultured in the medium with different concentrations of murine TNF-α for 48 hours, and saw its activity increase in low concentration (0.01 ng/mL, 0.1 ng/mL) compared to the negative control group but decrease in high concentration (10 ng/mL, 100 ng/mL). Meanwhile, when IκBα as an inhibitor of NF-κB was gene-introduced to the ST2 cell and over-expressed, the IκBα effect was reversed in the experimental group with high concentration (100 ng/mL) of TNF-α in the culture medium, and ALP activity increased by about 2.5 times that of the negative control group. When they analyzed mineralization after long-term (4 weeks) culturing, however, the mineralized area of the cell cultured in the medium with reduced TNF-α as well as whether it was over-expressed or not were not known48. Paula-Silva et al.49 cultured DPSCs in the medium containing TNF-α (10 ng/mL) for 24 hours and detected the expression of DPP, DSP, DMP-1, and OC. This means that TNF-α plays the role of inducing DPSCs to differentiate into odontoblast. All the results above show that mesenchymal stem cells induce osteoblastic/odontoblastic differentiation when TNF-α is added for a short time, but that the opposite result is obtained when TNF-α is added for long time.
5. TGF-β
As a prototypical member of the TGF-β superfamily, TGF-β is one of the cytokines existing the most in bone matrix; it plays two contrary roles: promoting early oste-oblastic differentiation and obstructing late osteoblastic differentiation and mineralization54,55. Many research studies reported that TGF-β obstructed BMP-induced osteoblastic differentiation56,57. According to de Gorter et al.45, however, the BMP-6-induced ALP activity decreased when the TGF-β receptor was silenced in murine MSCc through the kinase siRNA screen. Likewise, as a result of inducing osteoblastic differentiation for 16 days with the gene introduction of both BMP-6 and TGF-β in the medium, genes related to osteogenic differentiation, ALP activity, and mineralization increased much more compared to the experimental group, which only had BMP-6 for inducing osteoblastic differentiation. When they put TGF-β continuously for 16 days, mineralization was obstructed instead. This suggests that TGF-β can promote or suppress osteogenic differentiation according to the culturing condition, period, and duration.
6. Leptin
As 16 kDa protein encoded in the Ob gene, leptin is known to play the role of adjusting food intake. It is known to be secreted by osteoblast during bone formation and to obstruct bone resorption and promote bone mineralization by suppressing osteoclast activity33,34. Nonetheless, a report claims that leptin obstructs bone formation through the hypothalamic signaling system58. According to the research of Um et al.33, which studied the influence of leptin on dental stem cells, when DPSCs and PDLSCs were cultured in the medium containing leptin, more mineralization was noted compared to the negative control group. In particular, the PDLSC experimental group was much more mineralized than the DPSC experimental group. The peculiar thing was that, when they put cells in the hydroxyapatite/tricalcium phosphate particle and implanted it into a nude mouse and observed it histomorphologically 12 weeks later, a similar cement structure formed from a similar cementoblast cell was observed in the PDLSC experimental group, and a similar dentin structure formed from a similar odontoblast cell - into which DPSCs differentiated - in the DPSC experimental group33. This shows that leptin plays the role of modulating the osteoblastic/odontoblastic differentiation of mesenchymal stem cells.
III. Review and Conclusion
In the area of oral and maxillofacial surgery, the tissue engineering approach to the reconstruction of maxillofacial defect or dental regeneration is the most discussed study field nowadays. To apply this effectively to the clinic setting, pre-clinical and clinical studies are ongoing. As the representative protein, BMP is actively used for the reconstruction of maxillofacial defect but has a drawback, i.e., a relatively high amount of BMP should be used to gain the beneficial effects of bone regeneration because BMP obstructs the effect of bone regeneration by inducing the expression of noggin, which gives negative feedback inherently, and it only works for a short time when applied directly to bone defect45. While clinically applying recombinant protein directly to a defect has a limitation, i.e., it is effective for a short time only, genetic transformation using the gene introduction of mesenchymal stem cells has significance since the biologic characteristic of the cell with gene introduction can be vitalized continuously23. Moreover, the aforesaid research confirmed that, when genes having mutual synergy (ex. BMP6+VEGF, BMP6+TGF-β, etc.) were introduced together instead of single gene introduction, the modulation of osteoblastic/odontoblastic differentiation was more effective.
DPSCs, which are easy to get surgically compared to BMSCs, are mesenchymal stem cells and are considered to be used in the regenerative cell therapy of other organizations such as liver as well as dental tissue regeneration therapy because of their multi-lineage differentiation capability, i.e., they can differentiate into osteoblast, adipogenic cell, cartilage cell, etc25. According to previous research, DPSCs formed mineralized tissue such as BMSCs when inducing osteogenic differentiation but had lower effectiveness than BMSCs44. Meanwhile, differentiated tissues show more odontoblastic differentiations than osteoblastic differentiation2. Based on this, we can expect DPSCs to be introduced for more specific differentiation inducement in the oral and maxillofacial area.
There are viral vector and nonviral vector methods when the genes are introduced to mesenchymal stem cells. There are drawbacks, however, such as low effectiveness (nonviral), short expression period (nonviral and adenoviral), induced immune reaction (adenoviral), and insertional mutagenesis (retroviral)23. Because of this risk, the method of gaining similar effect to gene introduction using recombinant protein is currently used, but it also has a limitation, i.e., it works only for a short time, and high concentrations are difficult to achieved5. For future clinical application, safer, biocompatible, more efficient, and effective vectors should be developed and can be considered for use together with recombinant protein. This study is significant since it reviewed and organized literature on the modulation of osteoblastic/odontoblastic differentiation through gene introduction to mesenchymal stem cells. In the future, there should be more research studies until tissue engineering using mesenchymal stem cells through gene introduction can be applied clinically to oral and maxillofacial reconstruction.
 XML Download
XML Download