

 PDF
PDF ePub
ePub Citation
Citation Print
Print


The AmpC β-lactamases produced by Enterobacteriaceae are clinically relevant enzymes that can hydrolyze penicillins, cephamycins such as cefoxitin, as well as broad-spectrum cephalosporins such as ceftazidime or cefotaxime. Moreover, penicillinase inhibitors are ineffective against these enzymes. The cephalosporinases encoded in the chromosome of group 3 Enterobacteriaceae were originally present only in Enterobacter spp., Morganella spp., Citrobacter spp., and a number of other species. Following the acquisition of ampC genes via transmissible plasmids, cephalosporinases began to appear in Escherichia coli and Klebsiella pneumoniae as well [1]. Distinguishing between acquired ampC and chromosomal ampC is possible by gene amplification only. Plasmid-mediated AmpC enzyme production is less common than production of extended-spectrum β-lactamases, and its detection remains difficult in the presence of other enzymes, leading to an underestimation of this resistance mechanism [2]. We identified K. pneumoniae strains that exhibited an AmpC phenotype in the absence of PCR-detectable ampC genes. Furthermore, we used genome sequencing and identified other β-lactamase genes that could possibly explain the resistance phenotype of these strains.
A total of 257 K. pneumoniae strains were collected from 17 hospitals in Korea. No ethical clearance was required for this study since patient data were not included and the source of the strains was anonymous. The strains were collected in 2014 and were stored at −80℃ until use. Before use, the strains were subcultured on MacConkey agar and were deposited in the central laboratory (Seoul St. Mary's Hospital). Of the total isolates, 39 were intermediately resistant or resistant to cefoxitin, and 31 of these expressed a plasmid-mediated DHA-type AmpC β-lactamase; this enzyme confers resistance to cephamycins and oxyimino-cephalosporins and is most likely characterized by inducible production. Phenotypic detection of AmpC was performed using a modified disk-based method, with boronic acid as an AmpC inhibitor [3]. Briefly, the isolates were tested using a standard cefoxitin disk (30 µg) and a cefoxitin disk supplemented with 400 µg of phenylboronic acid (Sigma-Aldrich, Saint-Quentin Fallavier, France). Isolates were considered AmpC-positive if the zone of inhibition surrounding the phenylboronic acid-cefoxitin disk was ≥5 mm larger than that surrounding the cefoxitin disk.
The isolates were also tested using a multiplex PCR assay that identifies plasmid-mediated ampC genes [4]. Five ampC-PCR-negative strains exhibiting the AmpC phenotype were subjected to further phenotypic characterization and whole genome sequencing (WGS) for detailed analyses of resistance gene content. The antimicrobial susceptibilities of the strains were tested using the VITEK2 instrument (bioMérieux, Marcy-l'Étoile, France). DNA was extracted using the Mo Bio kit (QIAGEN, Hilden, Germany) and sequenced using the Illumina MiSeq system with a 2×150 bp approach and a minimum coverage of 100×. The reads were assembled using A5-miseq, and the scaffolding was performed using SSPACE. The ResFinder online tool (https://cge.cbs.dtu.dk/services/ResFinder/) was used to assess the presence of (acquired) antimicrobial resistance genes, while the BioNumerics 7 software (Applied Maths, Sint-Martens-Latem, Belgium) was used to define microbial multi locus sequence types (MLST). The same tool was used to assess mutations in the ompK35 and ompK36 genes, which encode porins important for antibiotic uptake; mutation of these genes can lead to impermeability to several types of β-lactam drugs [5].
The five ampC-PCR-negative K. pneumoniae strains (all originating from urine samples and collected in different cities in Korea) belonged to four different sequence types (ST) (Table 1). Phenotypic characterization showed that all strains, except for 1606284, were resistant to cefoxitin; all strains were resistant to ampicillin/sulbactam and ticarcillin/clavulanic acid; and all but two strains (1606283 and 1606285) were resistant to piperacillin/tazobactam, according to the EUCAST breakpoints (Table 2). As the ResFinder results confirmed the absence of AmpC-encoding genes, we investigated which other genes could produce an AmpC-like phenotype.
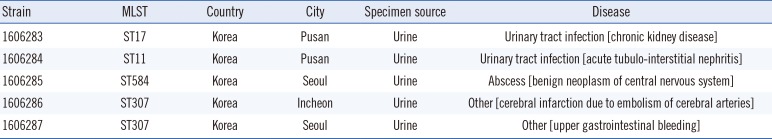
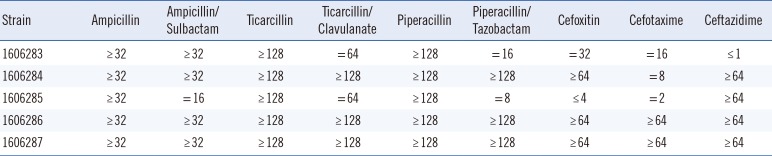
Table 1
Multilocus sequence type (MLST) and metadata of the five Klebsiella pneumoniae strains subjected to whole genome sequencing
![]()
Table 2
Minimum inhibitory concentrations in micrograms per millilitre of the five Klebsiealla pneumoniae strains
![]()
The enzymatic mechanisms conferring resistance or decreased susceptibility to β-lactamase inhibitors and cephamycins in K. pneumoniae include plasmid-mediated AmpC-type β-lactamase (p-AmpC) [1]; hyper-production of plasmid-mediated class A β-lactamases, such as TEM-1 and SHV-1 [67]; production of inhibitor-resistant TEM (IRT) β-lactamases [89]; plasmid-mediated OXA-type β-lactamase, such as OXA-1 [10]; and complex mutant TEMs (CMT), which are enzymes that combine IRT- and extended-spectrum β-lactamase (ESBL)-type substitutions [11]. ResFinder analyses identified at least one gene in each strain that could fully or partially explain the presence of a resistance phenotype in an ampC-PCR-negative background (Table 3).
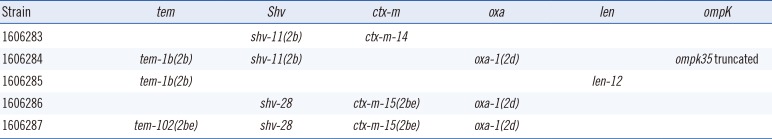
Table 3
The bla genes conferring resistance to β-lactams identified by ResFinder and BioNumerics analysis
![]()
ResFinder showed the presence of the blaOXA-1 gene in three of five strains (1606284, 1606286, and 1606287); two of these strains also contained blaCTX-M-15 and blaSHV-28, and the third strain also possessed blaTEM-1b and blaSHV-11. These three strains were associated with high minimum inhibitory concentrations (MICs) for both penicillin plus β-lactamase inhibitor combinations tested and for cefoxitin. Strain 1606283 carried blaCTX-M-14, encoding an ESBL able to confer decreased susceptibility to β-lactamase inhibitors; this strain demonstrated resistance to sulbactam and clavulanate and intermediate resistance to tazobactam. These results and the high cefoxitin MICs suggested impermeability to cephamycins. BioNumerics analysis of the ompK genes did not reveal any genetic mutations leading to frameshifts resulting in a truncated protein. However, we cannot exclude the presence of mutations leading to altered functionality or limited gene expression. Strain 1606285 carried blaTEM-1B together with blaLEN-12; overexpression of blaTEM-1B can decrease the susceptibility to combined treatment with penicillins and β-lactamase inhibitors. Overexpression of blaTEM-1 has been associated with promoters p3, pa/pb, p4, and p5 [12]; we observed that both strains carrying the tem-1b gene harbored the p3 promoter upstream of the β-lactamase gene. Finally, analysis of the ompK genes revealed that only the ompK35 in strain 1606284 contained a deletion that resulted in a frameshift and, consequently, a truncated protein.
In this pilot study, we show that discrepancies between resistance phenotypes and PCR results can be elucidated and possibly corrected using genomic data. Although these results are not unexpected, our analyses demonstrate that genomics data could be a valuable addition to PCR-based targeted gene testing. We have not included functional studies to prove that the new genes identified are causal of the phenotype because this was beyond the scope of the study. Furthermore, we assume that previously reported associations between phenotypes and the presence of certain genes (such as blaCTX-m-14, blaTEM-1b, and blaOXA-1) are correct and reliable [13]. Although PCR analysis is preferable in many settings because of the high cost and complexity of genomic analysis, we recommend that national reference laboratories adopt the pipeline proposed in our current analysis.
 XML Download
XML Download