

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Mycobacterium tuberculosis (MTB), an obligate pathogenic bacterial species that causes tuberculosis (TB), is a highly transmissible agent with significant mortality and morbidity. It has long been recognized that elderly people are more vulnerable to TB, and the TB treatment outcomes in elderly adults are often relatively poor owing to delayed diagnosis, more drug-related adverse effects, frequent co-morbidity, and overarching poverty [1]. Thus, targeted efforts to improve TB detection and treatment prognosis in the elderly population are needed to achieve global TB control [2].
Currently, the reference method for determining multidrug-resistantb (MDR) or extensively drug-resistant (XDR) TB is culture-based drug susceptibility testing (DST). However, this method is labor-intensive, time-consuming (taking weeks to months to obtain results), expensive, and technically challenging, especially in resource-limited countries [3]. Moreover, this methodology requires handling viable and potentially infectious cultures, specialized laboratory facilities, expensive consumables, and instrument maintenance [4]. There are currently several commercially available molecular assays to rapidly and reliably detect common mutations related to isoniazid (INH) and rifampin (RIF) resistance as potential alternatives to phenotypic DST, including the GenoType MTBDRplus line probe (LPA) [5] and GeneXpert MTB/RIF assays [6]. However, the absence of wild-type bands related to a specific gene on the LPA interpreted as resistant to the test drug may not be supported by a phenotypic assay [7]. Furthermore, one of the main current limitations of nucleic acid-based assays is that they do not accommodate all the mutations conferring resistance to anti-TB drugs.
Routinely, liquid acid-fast bacilli (AFB) cultures are sent to medical laboratories for phenotypic DST when they are culture-positive (usually one to three weeks after inoculation), and the results are reported after an additional two to three weeks according to breakpoints to determine if the cultures are susceptible or resistant to anti-TB drugs. Thus, clinically crucial information to initiate the most appropriate treatment is only available 4 to 6 weeks after a patient has been diagnosed with TB.
Next-generation sequencing (NGS) has emerged as a technique that can significantly contribute to TB control and treatment owing to its capacity to rapidly identify multiple gene targets associated with drug resistance-determining regions. Ion semiconductor sequencing is one of the available NGS platforms, which uses a small chip to detect released hydrogen ions emitted during DNA polymerization when deoxynucleotide triphosphates are incorporated into a DNA strand growing on a semiconductor device [8]. Several studies have described the clinical utility of Ion Torrent sequencing to characterize known mutations and discover novel variants linked to molecular-based drug resistance [91011]. We examined the feasibility of full-length gene analysis using ion semiconductor NGS with the Ion AmpliSeq™ TB panel (Life Technologies, Carlsbad, CA, USA) through comparison with the results of conventional phenotypic DST in 30 MDR-TB isolates using eight drug resistance-related genes.
Go to : 
METHODS
1. Sample preparation
The study protocol was approved by the Institutional Review Board of The Catholic University of Korea (DC16EISI0099). A total of 30 pure MDR-TB isolates were randomly selected and provided by the Korean Institute of Tuberculosis, Cheongju, Korea. Samples were requested for routine MTB culture and phenotypic DST from January to December 2016. Remaining samples revealing RIF and INH resistance in the phenotypic DST were sub-cultured again. Individual MTB colonies grown on the slants of 3% Ogawa solid medium within eight weeks in an incubator at 37℃ were carefully isolated. Mycobacterial genomic DNA was extracted from the pure MDR-TB isolates using a QIAamp DNA Mini Kit (Qiagen, Hamburg, Germany) according to the manufacturer's protocol. Purified genomic DNA was stored at −20℃ until analysis.
2. Phenotypic DST
Phenotypic drug resistance for first- and second-line drugs was determined using conventional DST (absolute concentration method with Löwenstein-Jensen medium) at the Korean Institute of Tuberculosis. The critical concentrations for each anti-TB drug tested were as follows: first-line drugs INH 0.2 µg/mL, RIF 40 µg/mL, ethambutol (EMB) 2.0 µg/mL, and second-line drugs streptomycin (SM) 10 µg/mL, amikacin (AMK) 30 µg/mL, kanamycin (KM) 30 µg/mL, and fluoroquinolones (FQs), including ofloxacin (OFX) 4.0 µg/mL, moxifloxacin (MXF) 2.0 µg/mL, and levofloxacin (LEV) 2.0 µg/mL. Pyrazinamide (PZA) susceptibility was estimated using a pyrazinamidase activity test. Besides INH and RIF resistance, the phenotypic DST results revealed 15 isolates resistant to EMB (50%), 10 isolates resistant to SM (33%), seven isolates resistant to PZA (23%), three isolates resistant to OFX (10%), two isolates resistant to LEV (7%), and one isolate resistant to MXF (3%); however, all 30 MDR-TB isolates were phenotypically susceptible to AMK and KM.
3. Ion semiconductor sequencing with the Ion AmpliSeq TB panel
Nucleic acid quality and quantity were assessed with the Broad-Range Qubit DNA kit and a Qubit 2.0 fluorometer (Life Technologies) followed by agarose gel electrophoresis. To identify gene variants correlated with drug resistance in genomic DNA extracted from the pure MDR-TB isolates, we generated AmpliSeq libraries using the Ion AmpliSeq Library Kit 2.0 and the Ion AmpliSeq TB Research Panel (Life Technologies). This panel amplifies 109 amplicons (two pools) from coding sequences of eight genes related to MTB drug resistance: inhA, katG, rpoB, pncA, rpsL embB, eis, and gyrA.
4. PCR amplification
The MDR-related genes were amplified through multiplex PCRs using 10 ng of genomic DNA with a premixed primer pool and the Ion AmpliSeq HiFi master mix (Ion AmpliSeq Library Kit 2.0) at 99℃ for 2 minutes 18 times (99℃ for 15 seconds and 60℃ for 4 minutes), followed by a hold at 10℃. We treated the obtained PCR amplicons with 2 µL FuPa reagent to partially digest the primer sequences, and phosphorylated the amplicons at 50℃ for 10 minutes, followed by 55℃ for 10 minutes and 60℃ for 20 minutes. We ligated the amplicons to adapters with diluted bar codes from the Ion Xpress Barcode Adapters kit (Life Technologies) for 30 minutes at 22℃ and then 72℃ for 20 minutes. We purified adapter-ligated amplicons (library) using Agencourt AMPure XP reagent (Beckman Coulter, Brea, CA, USA). The library was quantified using a Tapestation High Sensitivity DNA Assay kit (Agilent Technologies Inc., Santa Clara, CA, USA) and an Ion Library Quantification kit (Life Technologies) for quantitative PCR.
5. Emulsion PCR
Clonal amplification of the bar-coded DNA library onto ion spheres (ISPs) was conducted using emulsion PCR, and the DNA of the ISPs was subsequently isolated using an Ion PI HIQ OT2 200 kit and Ion OneTouch ES (Life Technologies) according to the manufacturer's instructions. We determined the polyclonal percentages and quality of enriched, template-positive ISPs using an Ion Sphere Quality Control kit (Life Technologies). We then sequenced samples with polyclonal percentages <30% and enriched, template-positive ISPs >80% on the Ion Torrent PROTON system (Life Technologies).
6. Sequencing using Ion Torrent Ion PROTON
We loaded the enriched ISPs onto Ion PI Chip v3, using 30 samples on a chip per sequencing run with the Ion PI HIQ Sequencing 200 kit (Life Technologies). For sequencing, we used 520 flow runs that generated approximately 200-bp reads. Ion Torrent sequencing was performed using the Ion PI HIQ Sequencing 200 kit, following the standard protocol.
7. Bioinformatics analysis
The raw signal data from sequencing runs were automatically transferred from Ion Torrent Ion PROTON to the Torrent Server hosted in the Torrent Suite software to process the raw voltage semiconductor sequencing data into DNA base calls. We aligned the sequences to the MTB H37Rv reference genome (NC_000962.3), and the base calling was conducted using Torrent Suite. Variants were identified with the Ion Torrent Variant Caller plug-in and Ion Reporter software (Life Technologies), and coverage maps were generated using a coverage analysis plug-in. Following data analysis, single nucleotide substitutions, insertions, and deletions were annotated with Ion Reporter software (Life Technologies). We then defined annotations of variants using snpEff v4.1 with upstream, coding, and downstream region information [12]. Amino acid changes such as missense, nonsense, or frame-shift variants were further evaluated. In addition, the sequence data were visually confirmed with Integrative Genomics Viewer (http://software.broadinstitute.org/software/igv/), and sequence, alignment, or variant call error artifacts were discarded.
For the rapid diagnosis of drug-resistant TB using sequence-based diagnostic methods, we searched the TB Drug Resistance Mutation Database (https://tbdreamdb.ki.se/Info/), which is a comprehensive list of genetic polymorphisms associated with first- and second-line anti-TB drug resistance, especially the most common mutations found for the major groups of anti-TB drugs in clinical MTB isolates detected worldwide [13].
8. Statistical analyses
To evaluate the agreement between genotypic DST using the Ion Ampliseq TB panel and phenotypic DST, we calculated the inter-rater agreement statistic (kappa) and 95% confidence interval (CI) for kappa, and interpreted each according to Altman's guidelines: poor, <0.2; fair, 0.21–0.40; moderate, 0.41–0.60; good, 0.61–0.80; and very good, 0.81–1.00 [14]. We then conducted statistical analyses using MedCalc ver. 17.6 (MedCalc Software, Mariakerke, Belgium), considering P <0.05 as statistically significant.
Go to :
RESULTS
1. Phenotypic and genotypic DST
Table 1 summarizes the detected amino acid changes in the 30 MDR-TB isolates using Ion Torrent semiconductor sequencing with Ion Ampliseq TB panels for inhA, katG, rpoB, pncA, rpsL embB, eis, and gyrA and compares the genotypic and phenotypic DST results. Overall, we identified 17 novel variants in the isolates: six (p.Gly169Ser, p.Ala256Thr, p.Ser383Pro, p.Gln439Arg, p.Tyr597Cys, p.Thr625Ala) in katG, one (p.Tyr113Phe) in inhA, five (p.Val170Phe, p.Thr400Ala, p.Met434Val, p.Glu812Gly, p.Phe971Leu) in rpoB, two (p.Tyr319Asp and p.His1002Arg) in embB, and three (p.Cys14Gly, p.Asp63Ala, p.Gly162Ser) in pncA. Because the Ion Ampliseq TB panel only targets coding sequences for these eight genes, we could not analyze the presence of common promoter mutations of inhA [15], eis [16], and pncA [17].
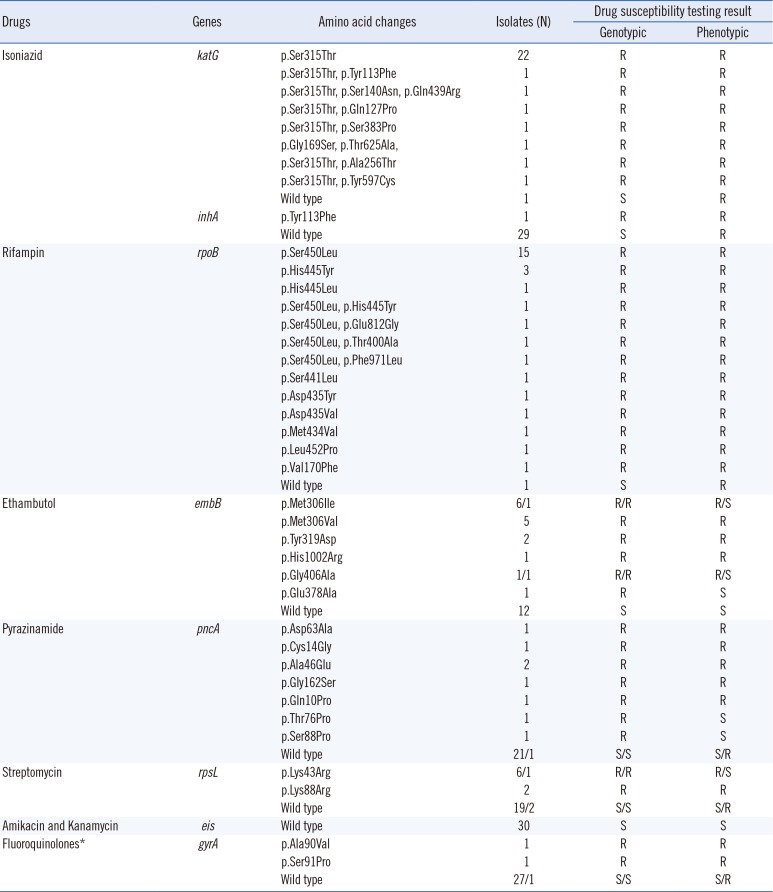
Table 1
Results of genotypic drug susceptibility testing by ion semiconductor sequencing in 30 multidrug-resistant Mycobacterium tuberculosis isolates
![]()
2. Genotypic DST using the Ion AmpliSeq TB panel as a predictor of phenotypic DST
The concordance between the Ion AmpliSeq TB and phenotypic DST results varied for each of the eight anti-TB drugs tested: for INH, RIF, EMB, PZA, SM, AMK+KM, and FQs, the concordance rates were 97%, 97%, 90%, 90%, 90%, 100%, and 97%, respectively, with agreement (kappa values) ranging from 0.734 to 0.800 (Table 2). The agreement rates for INH, RIF, and AMK+KM could not be estimated, because all isolates were phenotypically resistant to both INH and RIF, and all isolates were phenotypically and genotypically susceptible to AMK+KM. Overall, the genotypic DST results revealed “good” agreement with the phenotypic DST results for EMB, PZA, SM, and FQ. The overall major error rate, defined as phenotypic resistance with no mutation found, was 11% (4/35). Individual major error rates ranged from 0 to 33%: 0% for EMB (0/15), 14% for PZA (1/7), 20% for SM (2/10), and 33% for FQs (1/3). The overall major error rate defined as phenotypically susceptible with a high-confidence mutation related to drug resistance was 7% (6/85), whereas the individual major error rates ranged from 0 to 20%: 0% for FQ (0/27), 5% for SM (1/20), 9% for PZA (2/23), and 20% for EMB (3/15).
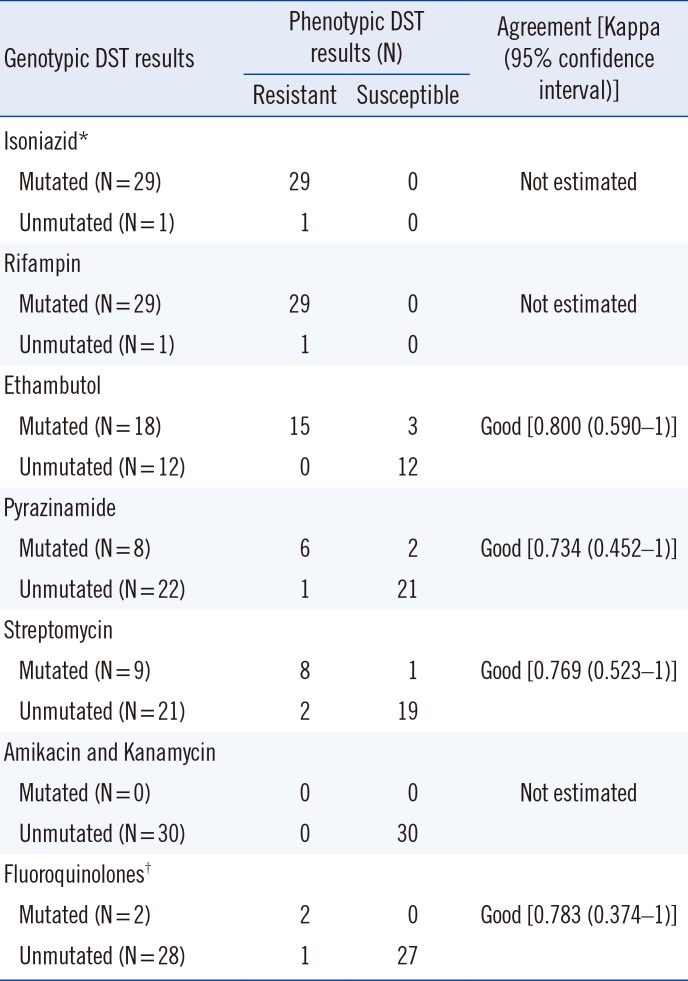
Table 2
Agreement between phenotypic and genotypic drug susceptibility testing by ion semiconductor sequencing
![]()
Go to :
DISCUSSION
We evaluated the feasibility of genotypic DST using ion semiconductor NGS with the Ion AmpliSeq TB panel by characterizing 30 MDR-TB strains. Currently available commercial molecular methods offer limited detection capabilities, particularly when uncommon or new amino acid changes are contained within common drug-resistance regions or when undetected amino acid mutations impact drug resistance [11]. This targeted NGS approach can characterize known mutations and facilitate the discovery of novel variants in the entire coding regions of eight full-length genes that have been previously implicated in MDR-TB resistance.
The katG gene encodes catalase peroxidase, an enzyme that converts INH into the active form. The majority of cases of INH resistance are associated with the katG codon corresponding to amino acid 315 (p.Ser315Thr) [18], although mutations in the ahpC and inhA promoter regions have also been reported to contribute to resistance [19]. Among the 30 strains we assessed, 28 (93%) contained the common amino acid change of serine to threonine at position 315 (p.Ser315Thr) that confers INH resistance. Novel double variants, namely p.Thr625Ala and p.Gly169Ser, in one MDR-TB strain were detected in a phenotypically INH-resistant strain.
Mutations within an 81-bp RIF-resistance determining region (RRDR) of rpoB are responsible for approximately 95% of all cases of RIF resistance in MTB strains [20]. Three of these mutations that yield substitutions at positions p.Asp435Val, p.His445Asp/Tyr, and p.Ser450Leu, respectively, constitute most of the mutations within this region. Of the 30 RIF-resistant strains characterized, 19 (63%) carried p.Ser450Leu, two (7%) contained an amino acid change at position 435, and five (17%) contained a mutation at position 445 of the encoded rpoB product. Interestingly, ion semiconductor sequencing revealed that five of the 29 rpoB-mutated strains contained a novel variant. We identified an uncommon amino acid change, p.Met434Val, at position 434 in the rpoB product, in which the most prevalent mutation was a change from methionine to isoleucine (p.Met434Ile) [21].
EMB is a first-line anti-TB agent used to prevent the emergence of drug resistance and is included in most MDR-TB regimens when susceptibility is demonstrated [22]. EMB interferes with mycobacterial cell wall synthesis and integrity by inhibiting arabinosyl transferases encoded by embC, embA, and embB [23]. The most common mutations in the EMB resistance-determining region (ERDR) are found in embB codons 306, 406, or 497; the remaining are located outside of these three positions between codons 296 and 426 or correspond to mutations in the embC-embA intergenic region [24]. Three of the MDR-TB isolates with embB mutations p.Met306Ile, p.Glu378Ala, or p.Gly406Ala were found to be phenotypically susceptible to EMB, in line with previous findings [25]. It has recently been proposed that the conventional phenotypic DST result for EMB should not be used in the design of individualized treatment for MDR-TB. The broth dilution method is more closely correlated with embB mutations than the proportion method in MDR-TB [26].
PZA, a synthetic derivative of nicotinamide, is used as a first-line drug, and PZA resistance is attributed to mutations in pncA encoding a pyrazinamidase [27]. However, these resistance-conferring mutations are variable and widely dispersed [28]. The difficulty in assessing PZA phenotypes and the sparse distribution of mutations along pncA highlight the useful application of NGS to assess mutations in this hypervariable gene.
SM, an aminocyclitol glycoside, interferes with translation proofreading, thereby inhibiting protein synthesis [29]. Various mutations in three genes, rrs, rpsL, and gidB, are involved in conferring resistance to SM [30]. We identified common mutations converting Lys to Arg in rpsL at codons 43 and 88 in eight of the 10 SM-resistant isolates, which is in line with previous reports [313233].
The main target of FQs is DNA gyrase, a type II topoisomerase composed of subunits A and B encoded by gyrA and gyrB, respectively [34]. Amino acid changes located within a short region of gyrA known as the quinolone-resistance determining region (QRDR) account for the majority of known FQ-resistant strains [35]. Two missense mutations, p.Ala90Val and p.Ser91Pro, were identified in the QRDR in two of three FQ-resistant isolates. Although gyrA and gyrB mutations can lead to ofloxacin resistance, there were no significant differences among strains with mutation locations in gyrA and gyrB, as well as levels of ofloxacin resistance [36].
From this perspective, ion semiconductor NGS with the Ion AmpliSeq TB panel has the potential to provide useful information several weeks before phenotypic DST results are available, even when the genotypic DST is conducted directly from culture isolates. In contrast to most Sanger sequencers, Ion Torrent sequencing does not require lasers, which are expensive and involve substantial maintenance, and typically require modified fluorescence-based, light-sensitive chemistry; thus, this platform offers a broader testing range at lower cost, without compromising assay performance or turn-around times in clinical settings [10]. We do not suggest that phenotypic DST should be replaced completely; however, for critical cases, there is clinical significance associated with the delay in obtaining broader DST results, to which genotypic DST can provide a complementary and more rapid result. In particular, genotypic DST can play a major role in determining resistance to the drugs EMB and PZA, for which phenotypic DST is known to be relatively less reliable [37]. Given that PZA is associated with a highly adverse effect of hepatotoxicity, especially in older patients, a genotypic DST for pncA may support phenotypic DST results and enable clinicians to manage and initiate anti-TB treatment earlier.
One of the main concerns related to performing full-gene NGS using only MDR-TB isolates is the discovery of novel mutations that may have no impact on drug susceptibility; conversely, this approach could reveal rare variants outside of known resistance sites that may be responsible for partial or complete resistance when accompanied by other mutations within either the same or other genes. The main limitation is that the Ion AmpliSeq TB panel is designed to cover the coding sequences of only eight genes. Thus, the panel should be revised to provide a primer that extends to promoter regions so as to cover the frequent promoter mutations associated with drug resistance, including inhA c.−15C>T [38], eis c.−14C>T, c.−12C>T, c.−10G>C/A [16], and pncA c.−11A>G, c−15A>C [1739]. In addition, the rrs gene encoding 16S rRNA should be included in the panel, which is associated with high-level resistance to second-line injectable drugs, such as AMK and KM, used to treat MDR-TB [40].
In conclusion, ion semiconductor NGS could provide the opportunity to detect reported or novel amino acid changes in the full coding regions of these eight genes. However, genotypic drug resistance profiles should be complemented and validated by subsequent phenotypic DST studies.
Go to :
 XML Download
XML Download