

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Klebsiella pneumoniae, a gram-negative bacillus, causes a wide array of infections resulting in severe morbidity and mortality. Isolates of this species have been reported to be resistant to almost all classes of antibiotics through progressive mutations in chromosomally encoded genes and acquisition of genes from mobile plasmids and integrons [1]. Carbapenems, widely used for the treatment of serious infections, are effective against K. pneumoniae, especially multidrug resistant (MDR) Enterobacteria producing high levels of AmpC cephalosporinases or extended spectrum β-lactamases (ESBLs). However, in recent years, widespread use of carbapenem has accelerated the growth of resistant strains in different regions [12]. The global spread of carbapenem-resistant K. pneumoniae (CRKP) has become a serious clinical challenge because of the limited treatment options.
The mechanisms underlying carbapenem resistance in K. pneumoniae species generally involve the production of carbapenemase and the loss or decreased expression of outer membrane proteins. Additionally, the production of AmpC and ESBL plays an important role in these resistance mechanisms [3]. KPC and NDM are the most notable carbapenemases associated with resistance mechanisms; the genes encoding these enzymes are found on transferable plasmids thus facilitating the spread of these resistance genes to different species worldwide. KPC-2 is the primary cause of carbapenem resistance among K. pneumoniae isolates in China [45]. However, recently, there have been increasing reports of K. pneumoniae harboring NDM-1 in several Chinese hospitals [6789]. More importantly, K. pneumoniae strains expressing carbapenemase or ESBL genes often co-harbor numerous drug-resistance determinants, such as quinolone and aminoglycoside resistance genes, which are partially responsible for MDR phenotypes; however, to date, little is known regarding the MDR mechanisms of CRKP. In addition, the high prevalence of ESBL-producing Enterobacteriaceae and the increasing use of carbapenems in our hospital necessitated an investigation of the epidemiology and molecular characterization of CRKP.
In this study, we isolated 78 CRKP strains from patients in our hospital from 2012 to 2015. We investigated the antimicrobial resistance phenotypes and underlying MDR mechanisms of the carbapenem-resistant strains.
METHODS
1. Bacterial strains
A total of 2,932 non-duplicated K. pneumoniae strains were isolated and identified from January 2012 to December 2015 by using the VITEK2 compact or VITEK MS systems (bioMerieux, Hazelwood, MO, USA) at the department of laboratory medicine of the First Affiliated Hospital of Chongqing Medical University, a major tertiary teaching hospital with 3,200 beds in Southwest China. Seventy-eight strains (2012, n=14; 2013, n=25; 2014, n=10; 2015, n=29) were resistant to at least one carbapenem on the basis of antimicrobial drug resistance determined by using the broth microdilution method [10]. These isolates were recovered from a variety of clinical specimens; urine (23/78,29.5%) was the most frequent source of CRKP, followed by respiratory tract (20/78, 25.6%), blood (9/78, 11.5%), bile (7/78, 9.0%), vaginal secretions (7/78, 9.0%), ascites (6/78, 7.7%), wound secretions (3/78, 3.8%), and abscess (3/78, 3.8%). The study was approved by the Biomedical Ethics Committee of First Affiliated Hospital of Chongqing Medical University.
2. Antimicrobial susceptibility testing
Antimicrobial susceptibility to ceftazidime (CAZ), ceftriaxone (CRO), cefepime (FEP), gentamicin (GM), tobramycin (TOB), ciprofloxacin (CIP), and levofloxacin (LEV) were determined initially by using AST GN13 cards on the VITEK2 compact system. The minimal inhibitory concentrations (MICs) of ertapenem (ETP), imipenem (IPM), and meropenem (MEM) were confirmed by using the reference broth microdilution method according to the CLSI methods [10]. The susceptibility testing results were interpreted according to the criteria recommended by the CLSI [11]. Escherichia coli ATCC 25922 was used as the quality control strain for susceptibility testing.
3. Determination of antibiotic resistance genes
PCR sequencing for the presence of β-lactamase genes, including carbapenemase-related genes (blaKPC, blaNDM, blaVIM, blaIMP, blaSME, and blaOXA-48) and other non-carbapenemase-related genes (blaCTX-M, blaTEM, blaSHV, and blaAmpC), was conducted by using previously described primers and conditions [12]. CRKP isolates were also tested for the presence of aminoglycoside resistant determinants (ARD, including aac(6′)-Ib, armA, and rmtB) and fluoroquinolone resistance determinants (QRD, including qnrA, qnrB, qnrS, and aac(6′)-Ib-cr). PCR analysis of the coding region of the ompK35 and ompK36 genes was performed by using previously described primers [13]. Integrons were screened and sequenced by PCR amplification of integrase genes int1, int2, and int3, as described previously [1415].
4. Restriction fragment length polymorphism (RFLP) analysis of gene cassette regions
To characterize integron variable region gene cassettes, the variable regions of class 1 integrons were further amplified to determine their gene cassette composition by RFLP analysis [16]. PCR products were subsequently digested with the RsaI and HinfI restriction enzymes. Integrons exhibiting the same RFLP patterns were considered to contain identical gene cassettes. Amplified DNA fragments of each distinct class 1 RFLP type integron were sequenced and analyzed by using the BLAST program (National Center for Biotechnology Information).
5. Transformation, conjugation, and plasmid analysis
The transfer experiment was carried out in mixed lysogeny broth cultures. Three CRKP strains co-producing blaNDM-1 and blaKPC-2 (K53, K55, and K65) served as the donors, while E. coli DH5α was used as the recipient strain. Transformants were selected on lysogeny broth agar plates containing 4 mg/L CRO. Similarly, the conjugation experiment was performed according to the previously described method [17] using rifampicin-resistant E. coli EC600 as the recipient strain on MacConkey agar plates in the presence of 8 mg/L CRO and 1,024 mg/L rifampicin. Transformants or transconjugants that grew on the selection medium were isolated, and their antimicrobial susceptibility was characterized by using the VITEK2 compact system (bioMerieux). Plasmid DNA extracted from donors, recipients, transformants, and transconjugants by using the alkaline lysis method was used as the template in PCR analyses. Plasmid replicons were determined by using the PCR-based replicon typing method [18].
6. Molecular epidemiology study
Pulsed-field gel electrophoresis (PFGE) was conducted to investigate the molecular epidemiology of the 78 CRKP isolates with XbaI-digested and genotyped chromosomal DNA. The genomic DNA restriction patterns of the isolates were analyzed and interpreted according to the previously proposed criteria [19]. In addition, to further determine whether clonal spread influenced the dissemination of carbapenemase-producing K. pneumoniae isolates in our hospital, multi-locus sequence typing (MLST) was performed by amplifying the internal fragments of seven K. pneumoniae housekeeping genes according to the MLST website (http://www.mlst.net).
7. Statistical analysis
All analyses were performed by using SPSS v.21.0 software (SPSS Inc., Chicago, IL, USA). Univariate analyses were performed separately for each of the variables. Categorical variables were calculated by using a chi-square test or Fisher's exact test as appropriate. Continuous variables were calculated by using Student t-test (normally distributed variables) or Wilcoxon rank-sum test (non-normally distributed variables) as appropriate. P<0.05 was considered statistically significant.
RESULTS
1. Increased rates of CRKP isolates, especially, carbapenemase-producing strains
Of the 2,932 non-duplicated K. pneumoniae strains, 78 were resistant to at least one carbapenem and were classified as CRKP. Interestingly, we found a statistically significant increase in CRKP frequency from 1.8% (14/765) in 2012 to 3.6% (29/802) in 2015 (P=0.031), as well as an increase of carbapenemase-producing strains from 28.6% (4/14) in 2012 to 69.0% (20/29) in 2015. Notably, while only six (7.7%) isolates were identified as NDM-1 producers from 2012 to 2014, the prevalence of NDM-1-producing K. pneumoniae increased dramatically to 20.5% (P<0.001) over the 2015.
2. Antimicrobial susceptibility testing
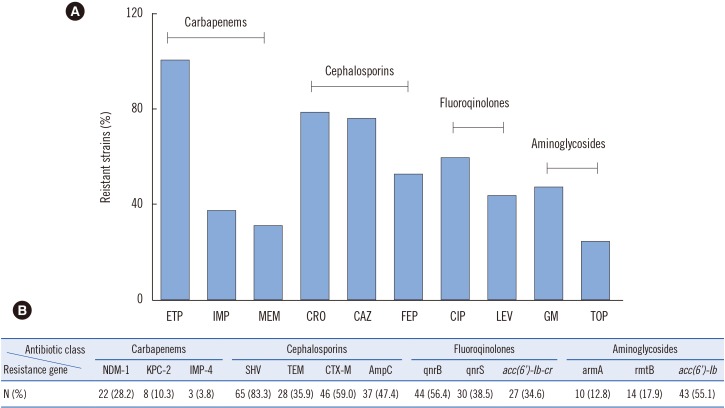
Of the 78 CRKP isolates, the ertapenem resistance rate was 100%, while 37.2% (29/78) and 30.8% (24/78) of the isolates were resistant to IPM and MEM, respectively (Fig. 1). In addition, the resistance rate to cephalosporins was relatively high. Specifically, 78.2%, 75.6%, and 52.5% of the isolates were resistant to CRO, CAZ, and FEP, respectively. The results of the fluoroquinolone testing showed that 46 (59.0%) and 34 (43.6%) isolates were resistant to CIP and LEV, respectively, while the aminoglycoside testing results showed that 37 (47.4%) isolates were resistant to GM, and 19 (24.4%) were resistant to TOB. Notably, 46.2% (36/78) of the CRKP isolates were classified as MDR as they were resistant to three or more classes of antimicrobials.
3. Resistance mechanism of the 78 CRKP isolates
Of the 78 CRKP isolates, 38.5% (30/78) harbored carbapenemase-encoding genes; 22 isolates possessed blaNDM-1, eight isolates contained blaKPC-2, and three isolates had blaIMP-4 (Table 1). It should be noted that the ETM, IPM, and MEM MIC ranges were dramatically higher in isolates possessing both blaNDM-1 and blaKPC-2 than those with only blaNDM-1. Moreover, carbapenemase-producing strains co-expressing blaESBL showed the highest MICs for ETM, IPM, and MEM. Unexpectedly, the MIC50 or MIC90 of three carbapenem agents were dramatically lower in the carbapenemase-producing isolates co-harboring ESBL and AmpC than in isolates positive only for ESBL; however, there was no statistically significant difference between the two groups (MIC50, P=0.268; MIC90, P=0.268).
The remaining 48 non-carbapenemase-producing isolates had a relatively high rate of ESBL genes (36/48, 75.0%) and AmpC enzymes (24/48, 50.0%). In addition to the production of ESBL or AmpC enzymes, deletion or mutation of porins such as OmpK35 and/or OmpK36 has been shown to be correlated with increased carbapenem MICs [20]. Surprisingly, only five (10.4%) of the 48 isolates were found to have defective porin genes, which differ completely from those observed in a multicenter surveillance study conducted in Korea [21]. Subsequent sequencing of the ompK35 and ompK36 genes of the remaining CRKP isolates (73/78) ruled out the occurrence of mutations or insertions, suggesting that these phenotypes were due to decreased expression, protein structure alteration, or inactivation of porins OmpK35 and OmpK36. Therefore, the production of ESBL or AmpC enzymes may have played an important role in carbapenem resistance in our hospital; differences in ESBL combinations or AmpC enzyme expression levels may explain the susceptibility profiles observed.
4. Prevalence of QRD and ARD genes in CRKP isolates
The prevalence of QRD genes in the 78 CRKP strains was 78.2% (61/78), with 44 strains carrying qnrB, 30 strains carrying qnrS, and 27 strains carrying aac(6′)-Ib-cr; 47.5% of the strains co-expressed at least two types of QRD genes. The prevalence of ARD genes was 65.4% (51/78 strains), and the genes aac(6′)-Ib, armA, and rmtB were detected in 55.1%, 12.8%, and 17.9% of the isolates, respectively; 17.9% of the strains co-expressed at least two types of ARD genes (Fig. 1).
5. Integron and gene cassette characterization
We found that nearly 66.7% (52/78) of the 78 isolates harbored the int1 gene, 45 were identified by amplification of the int1 cassette region, and the length of the amplicons varied from 0.3 kb to 2.6 kb. Gene cassette arrays were divided into 11 distinct types based on their RFLP profile. Moreover, 13 gene cassettes were detected in our study, of which the dfrA12-orfF genes were the most frequent (Fig. 2).
6. Plasmid analysis
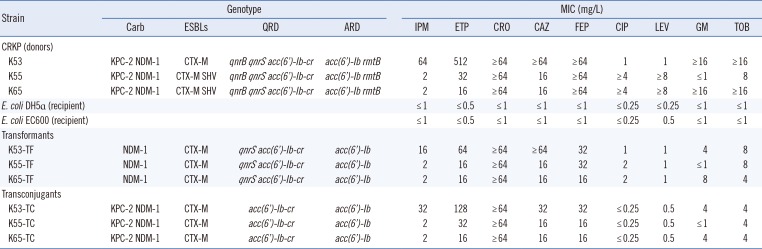
Carbapenem resistance was successfully transferred from three CRKP strains (K53, K55, and K65) to E. coli EC600 by conjugation and to E. coli DH5α by transformation. All E. coli transconjugants exhibited significantly reduced carbapenem susceptibility, with IPM and ETM MICs of 2–32 mg/L and 16–128 mg/L, respectively. In addition, the transconjugants exhibited MDR phenotypes similar to those of the clinical K. pneumoniae isolate donors. The transconjugants were also resistant to cephalosporins, but were susceptible to quinolones and aminoglycosides. Notably, three transconjugants simultaneously harbored blaNDM-1, blaKPC-2, blaCTX-M, acc(6′)-Ib-cr, and acc(6′)-Ib genes, while blaSHV, qnrB, qnrS, or rmtB were not detected in any of the transconjugants. The results of the transformation assays showed that three transformants were resistant to carbapenems and cephalosporins. Similar to the transconjugants, all transformants were susceptible to CIP and LEV; however, a number of transformants exhibited intermediate resistance to GM and TOB. Importantly, the transformation assays enabled the simultaneous transfer of blaNDM-1, blaCTX-M, qnrS, acc(6′)-Ib-cr, and acc(6′)-Ib genes in all of the transformants, while blaKPC-2, blaSHV, qnrB, and rmtB were not detected. In addition, plasmids from K55 and K65 (including those in the donors, transformants, and transconjugants) belonged to plasmid replicon type IncA/C, while the plasmids from K53 were untypeable (Table 2).
7. Molecular characterization
As shown in Fig. 2, 67 different XbaI patterns (1 to 67) were identified, indicating that the investigated isolates were epidemiologically unrelated. There were six patterns consisting of two different isolates that were closely related. However, these strains were collected from different wards. MLST demonstrated that sequence type (ST)11 was the most common among the carbapenemase-producing strains (6/30, 20.0%), followed by ST147 (4/30, 13.3%), ST290 (2/30, 6.7%), and ST2133 (2/30, 6.7%). The 22 blaNDM-1 positive strains were divided into 18 different ST patterns, of which the dominant clones, ST11 (3/22) and ST147 (3/22), exhibited diverse PFGE patterns and carried different antibiotic resistance genes. Notably, three isolates possessing both blaNDM-1 and blaKPC-2 belonged to ST1, ST147, and a novel type, ST1383. To the best of our knowledge, this is the first report on a K. pneumoniae ST1383 clone possessing blaNDM-1 and blaKPC-2 in China.
DISCUSSION
In the present study, the most plausible explanation for increased rates of CRKP isolates is the preferential use of carbapenem for the treatment of serious infections in our hospital, which has led to a gradual increase in carbapenem consumption in recent years. Increased usage may result in major selective pressure, thus promoting the progression of carbapenem resistance. As other studies have confirmed the high susceptibility rate of CRKP isolates to colistin and tigecycline [22], combination antibiotic treatment should be considered as the optimal treatment option for severely ill patients with serious infections caused by CRKP. In addition, the incidence of NDM-1 producers among the CRKP in our hospital (28.2%) was much higher than that previously reported in China [6789]. Notably, three NDM-1 isolates co-harbored the blaKPC-2 carbapenemase gene, which also contains other drug-resistance determinants. To the best of our knowledge, this is the first report on a MDR CRKP isolate co-harboring blaNDM-1, blaKPC-2, blaCTX-M, blaSHV, acc (6′)-Ib, rmtB, qnrB, and acc(6′)-Ib-cr. CRKP isolates carrying NDM-1 were more likely to be resistant to multiple antibiotics than CRKP strains lacking NDM-1 or other carbapenemases; NDM-1 was often accompanied by other genes encoding resistance to β-lactams, fluoroquinolones, or aminoglycosides, which was in line with a number of previous studies [2324].
Production of ESBL or AmpC enzymes has been demonstrated to play a role in K. pneumoniae resistance to broad-spectrum β-lactams and carbapenems [2]. In our study, we found 81.9% of the NDM-1 isolates co-expressed different ESBL and/or AmpC genes, exhibiting a complex β-lactamase background that was consistent with previous studies [2526]. The SHV-type ESBL gene was the most prevalent among the isolates, followed by the CTX-M-type, which is in agreement with two previous results [2728]. Furthermore, the identification of DHA-1 as the predominant AmpC in the CRKP isolates is similar to the recent findings in China [29].
The present study also demonstrated the qnrB was the most common gene and aac(6′)-Ib-cr was the least frequent gene identified in our hospital. Interestingly, aac(6′)-Ib-cr has been reported to be the most prevalent QRD gene in K. pneumoniae in many countries; these differences are probably due to regional variation in the prevalent resistant isolates [303132]. Most of the CIP and/or LEV resistant isolates identified in this study possessed at least one QRD gene; however, while several isolates were found to carry QRD genes, they exhibited susceptibility to CIP and LEV. Moreover, five isolates resistant to CIP and LEV were negative for QRD genes. Although qnr determinants alone may not confer resistance to quinolones, they do supplement other quinolone resistance mechanisms. Further studies will be required to examine additional resistance mechanisms mediating the high levels of fluoroquinolone resistance such as mutations in chromosomal gyrA and gyrC genes [33] or the presence of efflux pumps encoding genes such as qepA [34].
While many CRKP strains have been shown to be highly multiresistant, aminoglycosides may retain partial bactericidal activity against these isolates [35]. In our study, 52.6% (41/78) of the CRKP isolates were sensitive to at least one of the aminoglycosides tested. The high prevalence of aac(6′)-Ib is likely explained by the fact that the majority of CRKP isolates in our hospital possessed ESBLs; aac(6′)-Ib and ESBL genes are located on the same plasmid [36]. Notably, isolates carrying armA were more resistant to both GM and TOB than strains lacking this gene, suggesting that armA plays an important role in mediating higher aminoglycoside resistance rate.
Integrons, a diverse family of mobile elements, may also play an important role in the rapid evolution of MDR gram-negative pathogens by transferring antimicrobial resistance genes [14]. In the present study, class 1 integron was detected in 66.7% (52/78) of the CRKP isolates, and 13 different gene cassettes were observed in these isolates. The presence of integrons could be associated with resistance to trimethoprim, aminoglycosides, or fluoroquinolones, in agreement with a previous study [37]. Therefore, the wide distribution of integrons in CRKP isolates might pose a serious threat to the development of antimicrobial therapies.
Importantly, we found that all NDM-1 plasmids could be successfully transferred by both transformation and conjugation, whereas transfer of KPC-2 was successful only by conjugation. The most plausible explanation for this finding is that NDM-1 was carried on self-transmissible IncA/C or IncP plasmids, while KPC-2 was located either on non-self-transmissible plasmids or on the chromosome [38]. Moreover, the MICs of IPM and ETM against a number of transconjugants (K53-TC and K55-TC) were much higher than those against the transformants. The cephalosporin and aminoglycoside antimicrobial susceptibility patterns and resistance genes of the E. coli transconjugants were similar to those of the transformants. Interestingly, although both transformants and transconjugants were susceptible to quinolones, the MIC of CIP and LEV was 2- and 4-fold higher against the transformants, respectively, compared with the transconjugants, possibly due to the presence of qnrS in the E. coli transformants. Similar results were also found in a previous study [39].
It should be pointed out that none of the NDM-1 positive patients had a history of travel to any endemic area. Additionally, these NDM-1 isolates could not be attributed to the spread of a single clone, as determined by PFGE analysis; thus, all of the cases were sporadic. This local spread appears to be caused by different factors such as antibiotic abuse, poor infection control precautions, and environmental spread. Given the rapid spread of the NDM-1 carbapenemase in recent years, timely detection and aggressive control approaches are crucial to prevent its establishment as an endemic carbapenemase in our hospital.
In conclusion, our study demonstrates the high prevalence of NDM-1 genes encoding carbapenem resistance among K. pneumoniae isolates in Chongqing, China, as well as a high frequency of MDR determinants among these bacteria. Most importantly, this is the first report on a K. pneumoniae ST1383 clone carrying blaNDM-1 and blaKPC-2 in China and a MDR CRKP isolate co-harboring blaNDM-1, blaKPC-2, blaCTX-M, blaSHV, acc(6′)-Ib, rmtB, qnrB, and acc(6′)-Ib-cr. Because these isolates have limited treatment options, effective surveillance and strict infection control strategies should be implemented to prevent serious infections caused by CRKP in China.


 XML Download
XML Download