

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Hepatitis E virus (HEV) is the causative agent of epidemic and sporadic viral hepatitis, which has become a major public health concern in both developing and developed regions. Sporadic cases of zoonotic and food-borne infection have been reported in both HEV endemic and non-endemic areas [1]. Several investigations have shown that domestic pigs in industrialized areas are widely infected with HEV genotype 3, and autochthonous cases of hepatitis E have been reported [23]. The mortality rate of HEV infection ranges from 1% to 4%, and can be as high as 25% in pregnant women [4]. HEV is classified in the Hepeviridae family with at least four genotypes (1-4) of the virus infecting humans [5]. Genotypes 1 and 2 are restricted to humans, while genotypes 3 and 4 are zoonotic and responsible for autochthonous infections in humans [1]. HEV-3 is now considered an emerging pathogen, and is the most common genotype detected in both humans and swine in industrialized nations [267]. Several lines of evidence indicate the occurrence of the zoonotic transmission from pigs, wild boar, and deer to humans [89]. Furthermore, numerous reports have revealed the similarities in HEV-3 sequences detected in swine and humans from the same geographic are [1011]. In Europe, HEV-3 infections in pig farms are widespread with a prevalence of anti-HEV antibodies in pigs reaching 98%, and typically occur at the age of 2-6 months [12]. HEV infection in pig farms is also widespread in the USA [13]. However, to date, there is no information on the circulation of HEV in pigs and other animals in Bulgaria. HEV is not diagnosed routinely in pigs because the infection is often asymptomatic and is not associated with defects in growth [14]. This fact, together with sporadic cases of HEV infection occurring after the consumption of undercooked animal meat [151617], makes HEV a significant zoonotic disease that needs to be properly diagnosed.
HEV is a small, spherical, non-enveloped, single-stranded RNA virus [18]. The HEV genome has three open reading frames (ORFs). ORF2 encodes the capsid protein, which shows strong immunogenicity, and the antibodies generated in response to infection can efficaciously neutralize the virus [19]. Thus, the ORF2 capsid protein is an appropriate candidate for the serological diagnosis of HEV [202122].
Definitive diagnosis of HEV infection is based on detection of anti-HEV IgM antibodies or HEV RNA in serum samples. The presence of anti-HEV IgG is typically used to determine the seroprevalence in a population; however, both IgM and IgG antibodies are produced in people with acute HEV infection prior to the manifestation of clinical symptoms, which is potentially diagnostically relevant [23]. Therefore, patients presenting with infection symptoms will likely be positive for both anti-HEV IgM and IgG antibodies, although IgG is less indicative of an acute infection. Although several diagnostic kits for anti-HEV antibodies are available, they often provide contradictory results and are quite expensive [24]. Thus, the cost-effective production of recombinant proteins as diagnostic antigens is needed for the serological diagnosis of HEV. Although good expression has been achieved in both mammalian and bacterial cells, the process is quite expensive and cumbersome [25]. Therefore, plants may be a novel source for the cost-effective production of recombinant proteins useful for immunological studies. Plants have been explored as platforms for the production of diagnostic and therapeutic recombinant proteins owing to their scalability, safety, and their ability to perform eukaryotic post-translational modification [2526].
In this study, we developed a cost-effective and simple method for production of the HEV-3 ORF2 protein in Nicotiana benthamiana leaves, and demonstrated its usefulness for the serological detection of anti-HEV antibodies in both humans and swine. Transient expression in plant leaves enabled the efficient production of valuable proteins within one week. For this purpose, we used the Cowpea Mosaic Virus (CPMV)-based vector pEAQ-HT [27] for the expression of the HEV ORF2 protein in plant tissue. The expressed protein was recognized by both human and swine sera and can be used as a diagnostic reagent for the detection of anti-HEV IgG antibodies.
METHODS
1. Synthetic gene and plasmid construction
The nucleotide sequence of the swine genotype 3 HEV capsid protein (GenBank accession number DQ079627.1) lacking the first 109 amino acids (aa) from the N-terminus and 50 aa from the C-terminus (HEV 110-610) was synthesized by Life Technologies (Carlsbad, CA, USA). To maximize the levels of expression, the HEV ORF2 gene was codon-optimized for expression in N. benthamiana.
DNA fragments for insertion of the 6x His-tag at either the N- or C-terminus of HEV 110-610 was amplified by PCR. The primer pair HEV/C-end His-tag/Fw (AAATACCGGTATGGCTACTTCTCCT) and HEV/C-end His-tag/Rv (AGGCCTCGAGCTAATGATGGTGATGGTGATGAGCAAGAGCAGAGTGAGGAGCAAG) was used to clone the His-tag at the C-terminus, and the primer pair HEV/N-end His-tag/Fw (AAATACCGGTATGCATCACCATCACCATCATGCTACTTCTCCTGCTCCAGATACTGCT) and HEV/N-end His-tag/Rv (GGCCTCGAGCTAAGCAAGAG) was used for introducing a His-tag at the N-terminus. The PCR fragments were flanked by AgeI and XhoI restriction sites (New England Biolabs, Ipswich, MA, USA), which were used for the cloning of PCR fragments into the pEAQ-HT vector digested with the same enzymes. Finally, the plasmid DNA was purified and confirmed by PCR and sequencing. The E. coli XL1 blue strain was used for all cloning experiments.
2. Transformation of Agrobacterium tumefaciens
The constructs were transformed into the electrocompetent A. tumefaciens strain LBA4404. After electroporation at 2.5 kV, Super Optimal Broth with catabolite repression (SOC) medium was immediately added, and the cells were left to recover for 1 hr at 28℃. The cells were then plated on LB agar containing 50 µg/mL rifampicin and 50 µg/mL kanamycin.
3. Plant growth conditions
N. benthamiana plants were grown in glasshouses maintained at 25℃ and watered daily. Supplemental lighting was provided to maintain 16 hr of daylight in the winter months. Plants that were 3–4 weeks old were used for the transient expression experiments.
4. Agroinfiltration
Inoculated liquid cultures of A. tumefaciens strain LBA4404 were grown at 28℃ in a shaking incubator for 24 hr in Luria-Bertani (LB) medium containing rifampicin and kanamycin. N. benthamiana leaves were agroinfiltrated via a syringe at an optical density at 600 nm of 0.4.
5. Harvesting
Leaf tissue was harvested at six days post infiltration (dpi). Large-scale sampling was conducted by removing any large veins and non-infiltrated tissue, and recording the leaf sample weight.
6. Protein extraction and sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) analyses
Samples were extracted by adding 3 volumes of the extraction buffer (phosphate buffered saline, PBS, pH 7.4) with complete EDTA-free protease inhibitor cocktail tablets (Roche Diagnostics GmbH, Mannheim, Germany) and homogenized in a blender. Large cell debris was removed by squeezing the homogenate through one layer of Miracloth (Merck KGaA, Darmstadt, Germany). NuPAGE Bis-Tris Mini gels of 4–12% or 12% (w/v) acrylamide (Invitrogen, Carlsbad, CA, USA) and the protein pre-stained standard SeeBluePlus 2 (Invitrogen) or Spectra multicolor broad range protein ladder (TermoFisher scientific, Waltham, MA, USA) were used throughout the experiments.
7. Purification of recombinant HEV 110-610 His-tag protein
The recombinant protein from leaf extracts was purified using immobilized metal-anion chromatography (IMAC) on a Ni-NTA column, according to the manufacturer's instructions (Qiagen, Hilden, Germany). Briefly, infiltrated leaf tissue was blended with binding buffer containing 10mM imidazole at 3-times the fresh weight of the leaves (FWT). The extract was filtered through two layers of Miracloth, and after centrifugation, the clarified extracts were added onto a Qiagen Ni-NTA column. The column was washed with 20mM imidazole and was eluted with a total of 4 mL of elution buffer containing 250mM imidazole. Protein concentrations were determined by densitometry of the stained gels using known amounts of purified bovine serum albumin as a control and by measuring the absorbance at 280 nm. Both proteins were purified up to 100 µg/g FWT using IMAC and detected by using SDS-PAGE and western blot.
8. Western blot
The purified proteins were transferred from the SDS-PAGE gel onto a nitrocellulose membrane (Bio-Rad Laboratories Ltd, Hertfordshire, UK). Membranes were blocked with 5% (w/v) non-fat dried milk in PBS with 0.05% Tween-20 (v/v) (PBST) and incubated in the primary monoclonal antibody mouse anti-hepatitis E ORF2 antigen antibody (ab101124; Abcam, Cambridge, UK) diluted 1:3,000 at room temperature for 1 hr and washed with PBST. The bound antibody was detected with secondary anti-mouse antibody-horseradish peroxidase (HRP) (ThermoFisher Scientific) diluted 1:30,000. The emitted luminescence from the ECL detection reagents (GE Healthcare Life Sciences, Buckinghamshire, UK) was detected with the ImageQuant LAS 500 system (GE Healthcare Life Sciences). For western blot using human and swine serum as the primary antibodies, the membrane was incubated in human or swine serum (dilution 1:400), and bound antibodies were detected with secondary anti-human antibody-AP (Sigma-Aldrich, S.Louis, MO, USA) diluted 1:5,000 or secondary anti-pig antibody-AP (KPL, Gaithersburg, MD, USA) diluted 1:10,000, respectively. One-step NBT/BCIP substrate (ThermoFisher Scientific) was used. Western blot is universally accepted approving assay for detection of antibodies and it is considered a gold standard for affirmation of results. In this work, western blot was used to evaluate the quality of our in-house ELISA, by determining the rate of false-positives or false-negatives.
9. Human sera
All human serum samples, with informed consents from the patients, were provided by the National Centre of Infectious and Parasitic Diseases, Sofia, Bulgaria. The serum samples were tested with commercial kits from Dia.Pro (Dia.Pro Diagnostic Bioprobes Srl, Milan, Italy), which detects serum IgG and IgM. All human sera were positive for both antibodies.
10. Swine sera
Thirty serum samples were collected from apparently healthy pigs (six months old) using blood collected from the ear vein by veterinarians. Additional 15 serum samples were collected from the intracardiac clot of slaughtered pigs at slaughterhouses post-mortem. Sera were aliquoted and stored at −20℃ until use. All swine serum samples were donated by the Department of Animal Genetics, Faculty of Veterinary Medicine, Trakia University, Bulgaria.
11. In-house ELISA based on recombinant HEV 110-610 protein
Microtiter plates (Greiner 96-well flat bottom) were coated with 50 µL/well of serial dilutions of purified protein in duplicate wells (12.5 to 100 ng/well) in PBS (pH 7.4) and incubated overnight at 4℃. After three washes with PBST, plates were incubated with 200 µL/well of blocking solution (PBST–5% [w/v] dry milk) and incubated for 1 hr at room temperature. Serum diluted 1:100 in blocking buffer was then added.
Plates were washed again with PBST before the addition of 50 µL/well of an HRP-conjugated goat anti-human IgG or anti-swine secondary antibody diluted 1:10,000 (KPL) or 1:8,000 (ThermoFisher Scientific), respectively. After incubation with the secondary antibody, plate wells were washed three times before 50 µL/well of the substrate solution (o-phenylenediamine, Sigma-Aldrich) was added. Plates were incubated in the dark at room temperature from 10 to 20 min, and then the reaction was stopped by the addition of 50 µL/well of 1M H2SO4, and the plates were read at 492 nm in a plate reader Epoch Microplate Spectrophotometer (BioTek Instruments Inc., Winooski, VT, USA). Ten previously characterized negative human serum samples by using commercial kits from Dia.Pro (Dia.Pro Diagnostic Bioprobes Srl) and four negative swine serum samples by using porcine kit PrioCheck HEV Ab (ThermoFisher Scientific) were used as controls to determine the cut-off value for the human and swine anti-HEV IgG ELISA assay. The cut-off value was calculated as absorbance value of the test sample/absorbance value of the negative control or P/N of ≥2.5. Samples with absorbance values above the cut-off (P/N) were considered positive [20].
RESULTS
1. Protein expression and purification of swine genotype 3 HEV 110-610 capsid protein
A synthetic gene of the ORF2 of HEV lacking the first 109 aa at the N-terminus and the last 50 aa at the C-terminus was designed. The sequences were codon-optimized for the N. benthamiana genome to increase the yields of expressed protein [28]. The synthetic gene was used as template for insertion of a 6x His-tag at either the N- or C-terminus by the PCR DNA amplification method (see Methods). The resultant PCR products were cloned into pEAQ-HT and, after confirming the sequence of the insert, the constructs were transformed into A. tumefaciens, and N. benthamiana leaves were agroinfiltrated.
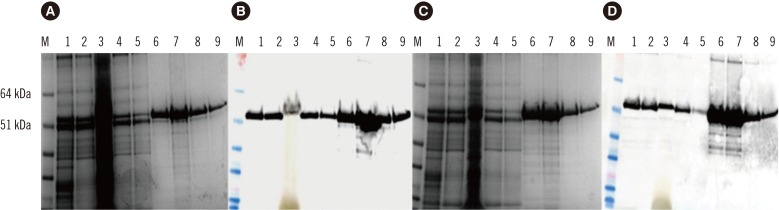
Plant leaves infiltrated with HEV 110-610 C-end expressed a large amount of a 54.6 kDa protein as revealed by SDS-PAGE followed by staining with Instant Blue (Fig. 1A). Western blot analysis using anti-HEV ORF2 antibody confirmed that the plants successfully produced HEV 110-610 C-end protein (Fig. 1B). The expressed HEV 110-610 with His-tag at the N-terminus was also analysed by SDS-PAGE (Fig. 1C) and confirmed with western blot (Fig. 1D). The 54.6 kDa band was absent in extracts from tissues infiltrated only with pEAQ-HT (data not shown).
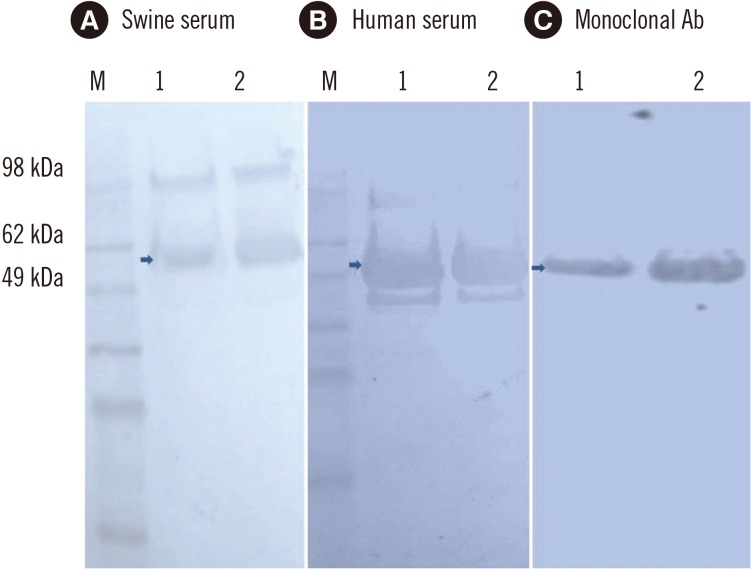
The purified protein was also run on SDS-PAGE gels and detected with anti-HEV IgG-positive swine sera (Fig. 2A) and with anti-HEV IgG-positive human sera (Fig. 2B). In addition, the samples were detected with anti-HEV ORF2 antibody as a control (Fig. 2C). Both the human and swine sera recognized a protein band of 54.6 kDa (arrows in Fig. 2) and some aggregates and degradation products.
2. ELISA based on the HEV 110-610 His-tag protein with human sera
The purified HEV 110-610 C-end and N-end His-tag proteins reacted similarly to human sera previously characterized as positive for anti-HEV antibodies. After obtaining the preliminary results with both proteins, we chose to perform all ELISA tests with the HEV 110-610 C-end His-tag protein. Five previously characterized negative human sera and five positive human sera were pooled and used for optimization of antigen concentration and serum dilution. Different concentrations of the HEV 110-610 recombinant protein were used. The optimal serum dilution was determined to be 1/100, and the antigen concentration was 50 ng/well (see Supplemental Data Fig. S1 and Fig. S2).
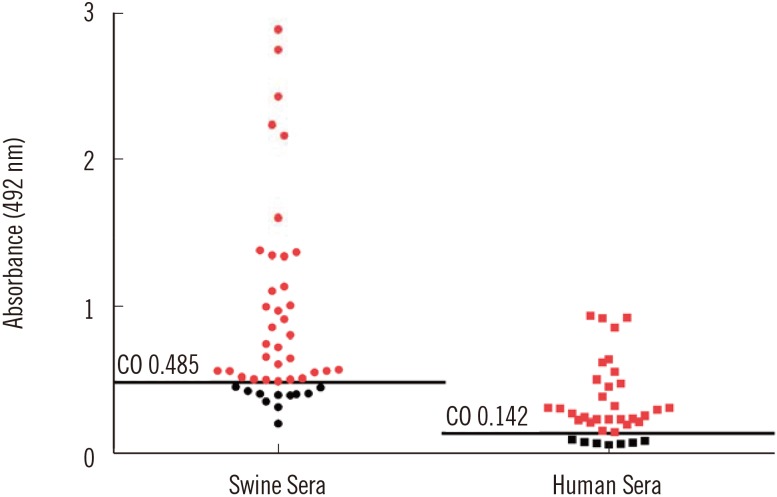
Thirty-six previously characterized positive human serum samples were tested by ELISA using the HEV 110-610 C-end His-tag protein as the antigen. The in-house ELISA showed anti-HEV IgG antibodies in 29 of the 36 human serum samples (Fig. 3).
Since western blot is more specific and confirmatory than ELISA, we also performed western blots with all of the human sera. All serum samples showed a positive signal on western blots. The sensitivity of the in-house ELISA assay was 80.5% compared with the commercial kit. From these data, we concluded that the plant-produced HEV 110-610-based ELISA test resulted in seven false-negative results (19.5%) (Fig. 4).
3. Detection of swine HEV antibodies
A total of 45 serum samples collected from healthy pigs in two Bulgarian farms and one slaughterhouse were tested for anti-HEV IgG antibodies. For specific HEV antibody detection in swine, we used both the plant-produced HEV 110-610-based ELISA and western blot. Four previously characterized negative swine sera and four positive swine sera were pooled and used for optimization of the antigen concentration and serum dilution. The optimal serum dilution was determined at 1/100, and the antigen concentration was 50 ng/well (see Supplemental Data Fig. S3).
The in-house ELISA showed antibodies in 34 of 45 pigs, indicating that 75.5% were positive for anti-HEV IgG antibodies (Fig. 3).
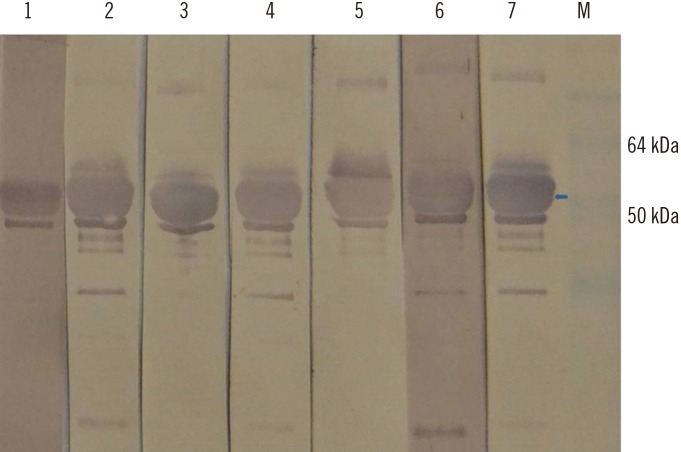
We then performed western blot with the 11 swine serum samples that scored negative by the ORF2-based ELISA. Four of these 11 samples tested positive by western blot (Fig. 5). This shows that the ELISA test resulted in four false-negative results or a rate of 8%.
DISCUSSION
We provide the first demonstration of the transient expression of truncated HEV 110-610 His-tagged protein in the plant N. benthamiana. The specificity of the expressed protein was confirmed by western blot using well-characterized monoclonal antibody, as well as human and pig sera. The optimal concentration of the antigen and serum dilution were assessed for an in-house ELISA for both swine and human sera. We demonstrated that the plant-produced truncated HEV ORF2 protein used as a coating antigen in our in-house ELISA was able to detect anti-HEV IgG antibodies in human sera with similar accuracy compared with that of the commercial kit. Since western blot is considered the gold standard, we used this method to confirm the ELISA-negative results. Anti-HEV IgG was detected in all human serum samples tested by the western blot. It is possible that some linear epitopes have been unmasked compared with the native antigen used in the ELISA. Furthermore, using the same method described herein, the recombinant plant ORF2 can also be used for the detection of serum IgM, which is considered to be more critical for the diagnosis of acute HEV infection (data not shown). Moreover, we could confirm the presence of the anti-HEV IgG antibodies in pigs in Bulgaria. Because pigs ordinarily are not tested for HEV infection, additional testing needs to be done to assess the seroprevalence of anti-HEV antibodies in Bulgarian farms.
Plant leaves infiltrated with pEAQ-HT HEV 110-610 with a His-tag at either the N- or C-terminus resulted in the expression of a 54.6 kDa protein at up to 100 µg/g of FWT. For instance, it has been reported that 1 mg of ORF2 protein can be produced per liter of E. coli culture [29]. Using the same protocol, 1 L of culture would produce enough protein to process 10,000 test samples, while 1 kg of plant tissue could produce enough protein to process 1,000,000 test samples at the yield described herein. This implies that one would need 100 L of bacterial culture to process the same number of samples as could be processed with the protein produced by 1 kg of plant tissue, which is much easier to obtain and much more cost-effective. If the production capacity is scaled up to levels used in the commercial production of plant-derived proteins (i.e., hundreds of kilos), then the final cost of the diagnostic reagent can quickly be lowered, improving its availability to those who need to administer the tests.
In conclusion, the HEV 110-610 recombinant protein can be produced in high yields at a relatively low cost in plant tissue, and could be used as a diagnostic antigen in the ELISA-based detection of anti-HEV antibodies in both human and swine sera.

 XML Download
XML Download