

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Methicillin-resistant Staphylococcus aureus (MRSA) is the most common pathogen causing hospital-acquired infections in many countries. Methicillin, an antibiotic active against penicillin-resistant Staphylococci, was introduced in 1959. Within two years, acquired methicillin resistance had emerged in S. aureus [1], and the resistance level steadily increased since. Currently, MRSA accounts for 60% of clinical S. aureus strains isolated from intensive care units in the United States [2], and this figure may be as high as 70-80% in most Asian countries [34]. MRSA exhibits multi-drug resistance, including resistance to all β-lactams. Therefore, antibiotics targeting MRSA are limited, but include vancomycin, linezolid, and daptomycin. However, resistance to these antibiotics has also been reported [567]. Thus, the development of new drugs or alternative therapies effective against MRSA is of great importance.
Natural products are now being re-assessed as antimicrobial agents [8], and the therapeutic potential of anti-MRSA phytochemicals has become increasingly recognized [9]. Moreover, one strategy to overcome the resistance mechanisms of various microorganisms is the use of drug combinations. Secondary metabolites from natural products are good candidates for combination therapies [10], and various natural anti-MRSA products have been identified [1112].
Phellinus linteus, P. baumii, and P. gilvus are medicinal mushrooms from the basidiomycetes Hymenochaetaceae family [13], a source of many antitumor or immunostimulatory polysaccharides, which have been utilized for centuries as folk medicines to treat various human diseases in several Asian countries. Of the wild mushrooms evaluated to date, P. baumii has been reported to exhibit anti-obesity [14], free radical scavenging [15], and hypoglycemic [16] properties. In addition, anti-platelet [17] and anti-inflammatory effects [18] have been reported, but antibacterial effects have never been explored. Herein, we evaluated the anti-bacterial effects of P. baumii ethyl acetate extract (PBEAE) against clinical isolates of MRSA as well as its synergistic effects when used in combination with β-lactams.
METHODS
1. Bacterial strains and characterization
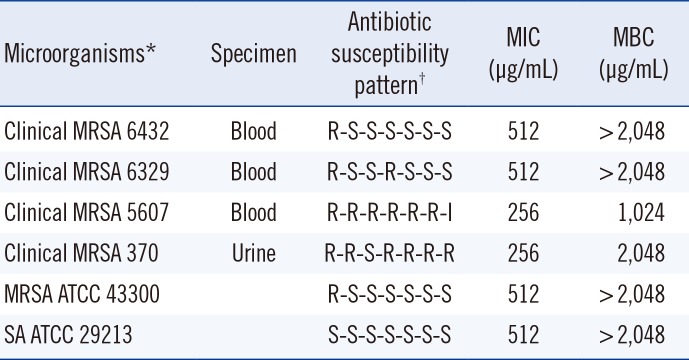
Four MRSA strains were isolated from blood or urine cultures from patients admitted to Chungbuk National University Hospital (Cheongju, Korea), in 2014. These strains showed different antibiotic susceptibility patterns. The S. aureus strains ATCC 29213 and 43300 were used as control methicillin-susceptible S. aureus (MSSA) and MRSA strains, respectively. MRSA was screened by disk diffusion for oxacillin-resistance, which was confirmed by using the penicillin binding protein (PBP)2a latex agglutination assay (MRSA-screen) (Denka Seiken Co., Tokyo, Japan) and PCR of mecA. Detection of β-lactamase in five MRSA and one MSSA strains was performed by nitrocefin disc (Becton Dickinson, Sparks, MD, USA). All isolates, including MSSA were positive for β-lactamase (Table 1).
2. Preparation of the PBEAE
P. baumii Pilat, identified by Professor Bong Sik Yun (mycologist), was collected from Geunsan Medicinal Herb Farm. Storage of the voucher specimen (PLPB-1001) and further extraction were performed as described previously [17]. Briefly, the dried fruiting body of P. baumii (10 kg) was extracted with methanol, filtered, concentrated, and partitioned with hexane and water. The aqueous portion was then subjected to ethyl acetate extraction to obtain the PBEAE, which was concentrated, dissolved in methanol, and then subjected to HPLC (Hitachi Co, Tokyo, Japan), nuclear magnetic resonance, and mass spectrometry analyses to identify and characterize the structure of the major active polyphenols such as davallialactone, interfungin A, and hypholomine B as described previously [15].
3. Antimicrobial agents and PBEAE
Eight antibiotics, including four β-lactams, were purchased. These were oxacillin (Sigma-Aldrich Co., St. Louis, MO, USA), cefazolin (Jongeun Dang Pharmceutical, Co., Seoul, Korea), cefepime (Sigma-Aldrich Co.), penicillin G (Keunhwa Pharmaceutical Co., Seoul, Korea), erythromycin (Sigma-Aldrich Co.), amikacin (Boryung Pharmaceutical Co., Seoul, Korea), ciprofloxacin (Bayer Korea Co., Seoul, Korea), and vancomycin (CJ Cheiljedang Pharmaceutical Co., Seoul, Korea). PBEAE was dissolved in dimethyl sulfoxide (Sigma-Aldrich Co.), while cefazolin and cefepime were dissolved in phosphate buffered saline (pH 6.0, 0.1 mol/L). Erythromycin was dissolved in 95% ethanol. Oxacillin, penicillin G, amikacin, ciprofloxacin, and vancomycin were dissolved in water. PBEAE and antimicrobial solutions were stored as aliquots at -70℃ prior to antimicrobial susceptibility testing.
4. Susceptibility testing of the PBEAE and antimicrobial agents
The minimal inhibitory concentrations (MICs) of the PBEAE and eight antibiotics (oxacillin, cefazolin, cefepime, penicillin, erythromycin, amikacin, ciprofloxacin, and vancomycin) and the minimal bactericidal concentration (MBC) of the PBEAE were determined by using the standard broth microdilution method with inocula of 5×105 colony forming unit (CFU)/mL following the CLSI guidelines [1920]. To evaluate the effects of the PBEAE in combination with antibiotics, the MICs were also measured by broth microdilution after the addition of one-half of the MIC (1/2 MIC) of the PBEAE. All experiments were performed in duplicate.
5. Bactericidal effects on MRSA and synergism
The synergistic anti-MRSA effects of the PBEAE and two β-lactams (cefazolin and oxacillin) were evaluated in a time-course bactericidal (time-killing) assay because of the dramatic decrease in MIC of the β-lactams when the extract was added. The interaction was defined as the log10CFU/mL increase in killing at 24 hr (ΔLC24) compared with that by the most active single drug, as follows: ΔLC24 ≥2 log10CFU/mL, synergy; ΔLC24=1-2 log10CFU/mL, additivity; ΔLC24=±1 log10CFU/mL, indifference; and ΔLC24 >-1 log10CFU/mL, antagonism [21]. The assays were performed in triplicate. The data are presented as mean±standard deviation.
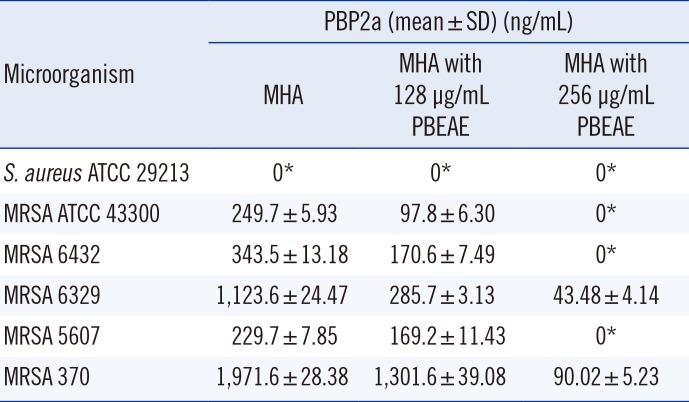
6. Measurement of PBP2a production by MRSA using an ELISA
To define the effect of the PBEAE on PBP2a production by MRSA, PBP2a was measured by using a sandwich ELISA. MRSA lysates were prepared from the isolates incubated on MHA with/without PBEAE (1/4 MIC=128 µg/mL; 1/2 MIC=256 µg/mL). The PBP2a concentration was corrected to the total protein concentration measured by using the BCA protein assay kit (Pierce, Rockford, IL, USA) in the MRSA lysates and analyzed. A mouse monoclonal antibody (mAb) against PBP2a 17A10 (1 µg/mL; Dinona, Seoul, Korea) was used as the capture antibody. Subsequently, PBP2a bound to the mAb was detected with horseradish peroxidase-conjugated mouse mAb against PBP2a 6G10 (Dinona) and 3,3',5,5'-tetraamethylbenzidine (TMB; Invitrogen, Carlsbad, CA, USA). The reaction was stopped after 10 min by the addition of 1.0 N sulfuric acid, and the absorbance was measured at 450 nm. The assays were performed in triplicate. The data are presented as mean±standard deviation.
RESULTS
1. Antibacterial activities of the PBEAE and combined effect with β-lactams on MRSA
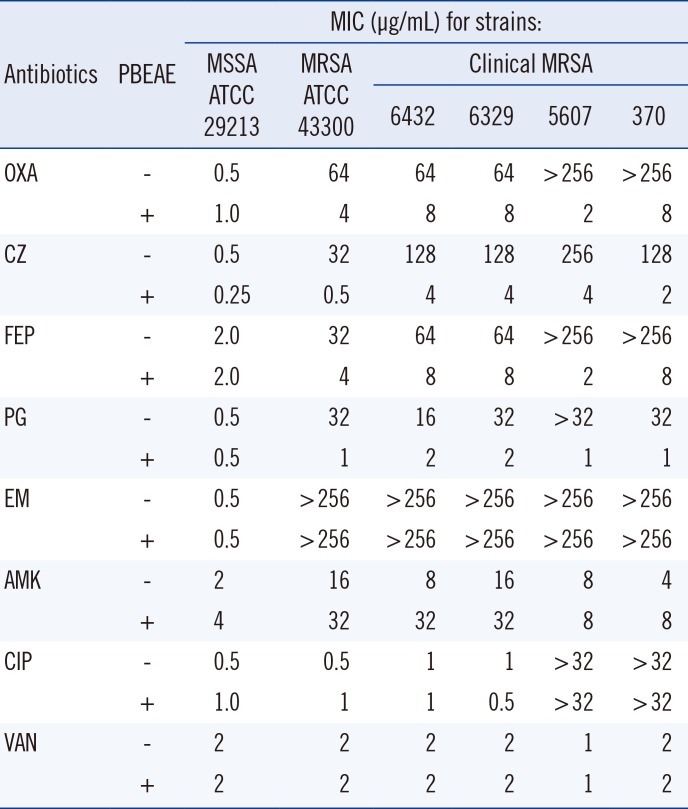
The PBEAE MIC and MBC against MRSA were 256-512 µg/mL and ≥1,024 µg/mL, respectively (Table 1). The combined effects of the PBEAE and various antibiotics against five MRSA and one MSSA are presented in Table 2. The PBEAE significantly reduced the MICs of various β-lactams (against MRSA strains) by 8-128 folds but had little or no effect on the activities of non-β-lactams.
2. Bactericidal synergistic effects of the PBEAE with β-lactams on MRSA
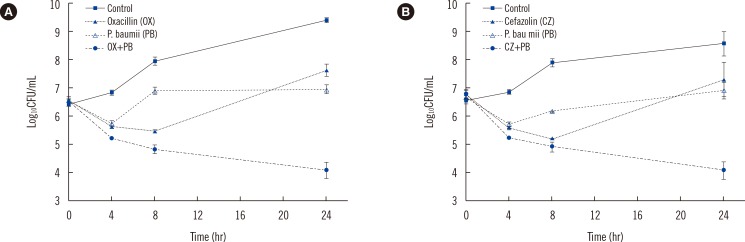
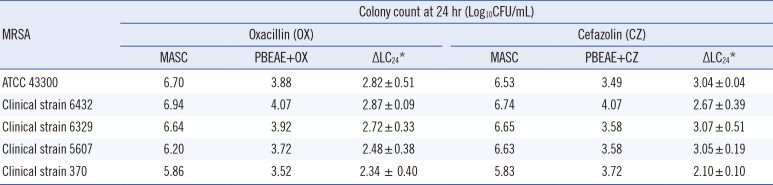
In the time-killing assay of a clinical MRSA strain (6432), the PBEAE alone displayed maximal killing effects at 8 hr followed by regrowth over 24 hr. Oxacillin or cefazolin alone displayed maximal killing effects at 4 hr followed by regrowth over 24 hr. However, PBEAE/oxacillin or cefazolin combination displayed maximal killing effects at 24 hr (Fig. 1). For all five MRSA strains, the combination of PBEAE/oxacillin or cefazolin increased the killing effect by >2 log10 CFU/mL at 24 hr, compared with oxacillin/cefazolin (the most active drug) or the PBEAE alone (Table 3), which demonstrated that the PBEAE had a bactericidal synergistic effect with β-lactams against MRSA.
3. Effect of the PBEAE on PBP2a production by MRSA
To identify the mechanism involved in the synergistic bactericidal effect of the PBEAE with β-lactams, the production of PBP2a by MRSA grown on MHA with or without the PBEAE was measured by ELISA. PBP2a production in MRSA grown on MHA with 1/4 MIC of the PBEAE was reduced to 25.5-73.66% of that of MRSA grown on MHA without the PBEAE. In MRSA isolates grown on MHA with 1/2 MIC of the PBEAE, PBP2a production was even reduced to 0-4.56% of that without the PBEAE (Table 4). The dose-dependent reduction in PBP2a production by MRSA suggests that the inhibition of PBP2a production in MRSA by the PBEAE is the major synergistic mechanism.
DISCUSSION
Of the medicinal mushrooms P. linteus, P. baumii, and P. gilvus, the antibacterial activities of P. linteus have been evaluated [22], whereas those of P. baumii have not. We first evaluated the in vitro antibacterial activities of PBEAE against MSSA (ATCC 29213), MRSA (ATCC 43300), and four clinical strains of MRSA. The PBEAE MIC and MBC were 256-512 µg/mL and ≥1,024 µg/mL, respectively, against MSSA and MRSA (Table 1). The PBEAE alone exhibited moderate to weak activity against MSSA and MRSA, being much weaker than the anti-staphylococcal antibiotics normally used to kill MSSA and MRSA (Table 2).
Various natural products exhibit not only anti-staphylococcal effects but also drug-synergistic antibacterial effects on MSSA and MRSA [911122223]. The combined effects of the PBEAE and various antibiotics against five MRSA and one MSSA are shown in Table 2. The PBEAE significantly reduced the MICs of various β-lactams (against MRSA strains) by 8-128 folds, but had little or no effect on the activities of non-β-lactams. The MICs of oxacillin and cefazolin (anti-staphylococcal β-lactams) were decreased to 8-128 folds and 32-64 folds, respectively. In addition, the PBEAE had no effect on MSSA, which does not produce PBP2a. These results show that the PBEAE inhibited the expression of PBP2a in MRSA, but not of other PBPs in MSSA.
The synergistic effect of the PBEAE and β-lactams on MRSA was evident when the extract was used in combination with the β-lactams, oxacillin or cefazolin (Table 3 and Fig. 1). Time-killing assays showed that the PBEAE/oxacillin combination increased killing by >2 log10 CFU/mL at 24 hr, compared with oxacillin (the most active drug) or the PBEAE alone, for all five MRSA strains (Table 3). In addition, the PBEAE/cefazolin combination increased killing by >2 log10 CFU/mL at 24 hr, compared with cefazolin or the PBEAE alone, for all five strains of MRSA (Table 3).
β-lactam resistance in MRSA is caused by the PBP2a encoded by mecA. PBP2a has low affinity for all β-lactams and PBP2a-producing MRSA are resistant to such antibiotics [2425]. There are two possible mechanisms of inhibition of PBP2a by the PBEAE. The PBEAE could inhibit PBP2a activity or inhibit its production. In this study, the production of PBP2a in MRSA grown on MHA with 1/4 MIC of the PBEAE was reduced to 25.5-73.66% of that of MRSA grown on MHA without the PBEAE. In MRSA isolates grown on MHA with 1/2 MIC of the PBEAE, PBP2a was further reduced to 0-4.56% of that without the PBEAE (Table 4). The PBEAE dose-dependent reduction in PBP2a production suggests that the PBEAE inhibited the production of PBP2a in MRSA, and this may be the major synergistic mechanism between β-lactams and the PBEAE. Nicolson et al. [26] showed that the diterpene derivative 416 potentiated the activity of methicillin by significantly reducing the expression of PBP2a. Meanwhile, Shiota et al. [12] reported that polyphenols (Corilagin and tellimagrandin I) isolated from Arctostaphylos uvaursi and Rosa canina, respectively, reduced the MICs of β-lactams toward MRSA through inhibition of PBP2a activity. P. baumii expresses several polyphenols, including davallialactone, interfungin-A, and hypholomine B [27]. The PBEAE polyphenols may inhibit PBP2a production in the MRSA strains. Therefore, combination therapy with the PBEAE polyphenols and β-lactam drugs may be useful as an alternative therapy for MRSA, similar to the combination of amoxicillin and clavulanic acid in β-lactamase-producing S. aureus. The PBEAE did not reduce MICs of β-lactams, including penicillin, in β-lactamase positive MSSA (S. aureus ATCC 29213). This result implies that the PBEAE does not inhibit β-lactamase production or activity (Table 2).
In conclusion, despite the fact that the PBEAE exerted a less potent antibacterial effect against MRSA compared with various antibiotics, the PBEAE significantly and synergistically potentiated MRSA killing by inhibition of PBP2a production when combined with β-lactams. This may be useful for the development of a combination therapy for patients infected with MRSA. Further studies, including larger numbers of clinical isolates, are warranted.
 XML Download
XML Download