

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Although 20-30% of the general population is asymptomatically colonized with Staphylococcus aureus, colonization increases the risk of S. aureus infection when the host immune system is compromised [1]. S. aureus infections vary from superficial wound infections to invasive diseases, such as deep abscesses, osteomyelitis, and bacteremia [2]. Methicillin-resistant S. aureus (MRSA) is of especially great concern because MRSA infection extends the length of hospital stay and increases antibiotic use, costs, and mortality [3].
Traditionally, MRSA strain typing was accomplished by pulsed-field gel electrophoresis (PFGE) [4]. However, the operational time of PFGE is at least 72 hr, the cost is relatively high, and the technique requires specialized training [5]. Sabat et al. [6] therefore designed a multiplex PCR scheme called multiple-locus variable-number tandem-repeat fingerprinting (MLVF), using 5 variable number tandem repeat (VNTR) loci. Studies have shown that MLVF can distinguish among MRSA strains and simultaneously provide results that parallel those of PFGE [5, 7, 8].
We aimed to investigate the usefulness of MLVF for typing S. aureus isolates of clinical significance. We also tried to modify the MLVF method described previously, in order to maximize the technical advantage of MLVF, in terms of time, cost, and simplicity, compared with PFGE.
METHODS
1. Bacterial isolates
Sixty-three hospital-acquired S. aureus isolates recovered in 2009 were selected according to Centers for Disease Control and Prevention (CDC)/National Healthcare Safety Network (NHSN) criteria [9]. Isolates that were regarded as contaminant or non-pathogenic were excluded. Among the 63 isolates, 15 were methicillin-susceptible, and 48 were methicillin-resistant. Most of the bacterial strains were isolated from blood culture and groups of 2 or 3 isolates that were considered epidemiologically related were chosen for analysis. The isolates were recovered from the following clinical sources: blood culture (N=43), ascitic fluid (N=2), cerebrospinal fluid (N=3), sputum (N=3), endotracheal aspiration (N=3), pus (N=2), T-cannula tip (N=2), wounds (N=2), throat (N=1), tissue (N=1), and drain (N=1).
2. DNA preparation
Bacterial isolates were subcultured in 5% sheep blood agar overnight at 37℃. Total genomic DNA was extracted from S. aureus isolates using the QIAamp DNA Mini Kit (Qiagen, Hilden, Germany), according to the manufacturer's instructions. Purified DNA was stored at -20℃.
3. MLVF typing
The multiplex PCR assay containing clfA, clfB, sdrCDE, spa, and sspA primers was performed as previously described by Sabat et al. [6], with modifications. For convenience, the AccuPower HotStart PCR PreMix kit (Bioneer Co., Daejeon, Korea) was used in the PCR reaction. The kit contained a mixture of 1 unit of HotStart DNA polymerase, 1× PCR buffer, 250 µM of each dNTP, and 1.5 mM MgCl2 in each reaction tube. A mixture of the following concentrations of each primer and 1 µL of template DNA (20-50 ng) was added to the reaction tube: 0.15 µM of clfA-F (forward) and clfA-R (reverse), 0.2 µM of clfB-F and clfB-R, 0.15 µM of sdrCDE-F and sdrCDE-R, 0.1 µM of spa-F and spa-R, and 0.3 µM of sspA-F and sspA-R. The thermal cycling was performed in a PTC-100 Thermal Cycler (MJ Research, Waltham, MA, USA) as follows: 94℃ for 5 min, followed by 30 cycles at 94℃ for 30 sec, 55℃ for 30 sec, 72℃ for 1 min, with a final extension at 72℃ for 7 min.
4. Visualization of PCR products
The amplified PCR products and 100-bp DNA ladder marker (Roche Applied Science, Indianapolis, IN, USA) were resolved by electrophoresis in a 2% agarose gel in 0.5× Tris-borate-EDTA (TBE) buffer at 50 V for 80 min using Mupid-2plus (Takara Bio Inc., Otsu, Japan). The gel image was saved as a TIFF file and analyzed using InfoQuestFP Software (Bio-Rad Laboratories Inc., Hercules, CA, USA) under the following tolerance settings: optimization, 0.5%, and position tolerance, 1.25% [5]. The band patterns were clustered into MLVF types using Dice's coefficient and the unweighted pair-group method with arithmetic mean (UPGMA). The effect of grouping similar MLVF patterns together into a smaller number of MLVF types was examined using cutoffs of >75% and >70% relatedness.
5. PFGE
Genomic DNA of the isolates was prepared, digested by restriction enzyme SmaI (Roche Diagnostics GmbH, Mannheim, Germany), and separated in a GenePath system (Bio-Rad Laboratories Inc.) according to the CDC PulseNet protocol with modification [10]. SmaI PFGE patterns were saved in TIFF files. The file was exported to the InfoQuestFP software (Bio-Rad Laboratories Inc.) and analyzed using the Dice coefficient-UPGMA. A dendrogram was generated to examine the relatedness of PFGE patterns for all study isolates, and cutoff levels of 80% and 75% were applied to this dendrogram.
6. Calculation of concordance
Simpson's index of diversity was calculated to measure the discriminatory power of the typing systems. This index indicates the probability that 2 strains sampled randomly from a population will belong to 2 different types [11]. The formulas for Simpson's index (D) and the confidence interval (CI) are presented in the following equations:
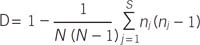
where N is the total number of strains in the sample population, S is the total number of types described, nj is the number of strains belonging to the jth type, and πj is the frequency nj/N.
To compare the 2 sets of results produced by PFGE and MLVF, we used 2 clustering comparison coefficients (adjusted Rand and Wallace) generated by a script from http://biomath.itqb.unl.pt/ClusterComp, as previously described [11]. We also evaluated the concordance of each pair of isolates as previously described [12]. All possible pairs of isolates were cross-classified on the basis of matched or mismatched types. The resulting 2×2 table was evaluated using the chi-square statistic, and the percentage of concordant cells was calculated.
RESULTS
1. Technical aspects of MLVF
Using a commercial kit containing a premixture of PCR reagents, we were able to perform MLVF more easily by simply transferring the template DNA and primers into the reaction tubes. This MLVF protocol, modified from a previous study [6], is sufficiently simple for use in clinical laboratories, and 18 isolates can be typed in a single working day if pure cultures of bacterial isolates are available. The hands-on time for 18 tests was approximately 3 hr on average (2 hr for DNA extraction, 30 min for preparation of PCR reaction, and another 30 min for MLVF pattern reading), with most of the time spent on the extraction of DNA. Experienced personnel would be able to analyze the MLVF patterns by computer software within 30 min.
2. Molecular typing by PFGE and MLVF
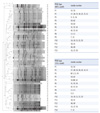
All 63 S. aureus clinical isolates could be typed by PFGE (Fig. 1) and MLVF (Fig. 2). Because defining clonality in the PFGE using a similarity value of 80% as a cutoff is considered the gold standard [13], we used 80% and 75% cutoff values for PFGE and 75% and 70% for MLVF. We also tried an 80% cutoff for MLVF; however, we found that some isolates were not grouped into the same type predicted by PFGE (data not shown). PFGE distinguished the isolates into 13 or 14 types and an additional 24 or 20 unique patterns at cutoff values of 80% and 75%, respectively (Fig. 1). Similarly, MLVF separated the 63 S. aureus isolates into 12 or 13 types and additional 16 or 12 unique patterns at cutoff values of 75% and 70%, respectively (Fig. 2). PFGE types P10 and P13 were identical to MLVF types M1 and M5, respectively. A very good correlation was observed between PFGE types P8, P5, and P6 and MLVF types M3, M4, and M9, respectively. In these types, only 1 or 2 isolates composing each MLVF type were different from the isolates composing the PFGE type. However, MLVF type M8, which was composed of 9 isolates at both 75% and 70% cutoff values, was divided into 5 different PFGE types, indicating that MLVF had a poorer discriminatory power than did PFGE.
3. Comparison of typing methods
To compare the discriminatory power of PFGE and MLVF, we determined the mean number of isolates per type and Simpson' s diversity index for all the isolates (Table 1). MLVF (75% cutoff) and PFGE (80% cutoff) yielded similar D-values of 0.954 (95% CI 0.933-0.974) and 0.972 (95% CI 0.957-0.988), respectively.
To compare the congruence between type assignments using PFGE and MLVF, we calculated adjusted Rand indices (Table 2) and Wallace coefficients (Table 3) for each cutoff value of PFGE and MLVF. The overall congruence between PFGE and MLVF was presented by adjusted Rand indices, ranging from 0.266 to 0.278. As expected from the Wallace coefficients, PFGE predicted MLVF type better than MLVF predicted PFGE type (Wallace coefficients, PFGE cutoff 80% vs. MLVF cutoff 75%, 0.389 vs. 0.233; PFGE cutoff 80% vs. MLVF cutoff 70%, 0.426 vs. 0.232; PFGE cutoff 75% vs. MLVF cutoff 75%, 0.359 vs. 0.256; PFGE cutoff 75% vs. MLVF cutoff 70%, 0.391 vs. 0.253).
Although the congruence between PFGE and MLVF in typing these clinical isolates was poor, notable agreement was found when the relationship between a pair of isolates was analyzed (Table 4). Cross-classification of the isolates, based on matched or mismatched schemes by PFGE (80% cutoff) and MLVF (75% cutoff), showed that the 2 typing systems were 91.0% concordant. Using different cutoffs, the PFGE and MLVF showed 88.3% to 90.1% concordance (data not shown). Any given pair of isolates distinguished by one method tended to be distinguished by the other.
DISCUSSION
Several studies based on VNTR schemes have been published for S. aureus strain typing [5-8, 14-16]. Most of the studies, however, were performed using selected MRSA isolates of certain limited PFGE types, which might introduce a selection bias. In this study, we chose clinically significant S. aureus isolates to evaluate the utility of MLVF, with respect to infection control in a hospital setting. Because PFGE is labor intensive, time consuming, and expensive, routine use of this procedure in a clinical laboratory is rather demanding. Therefore, we tried to determine whether PFGE could be replaced with the more "simplified" MLVF. We used a commercial premix PCR kit, instead of preparing a mixture of PCR reagents before amplification. After PCR amplification, we performed electrophoresis for only 80 min with a 12-cm length system, which can easily be utilized in clinical laboratories. Our modified method could save much time, compared to PFGE, which requires approximately 3 days (2 days for DNA agarose disc preparation, followed by 1 day of PFGE), as well as the previous MLVF procedure that utilized a master mix, which was prepared by adding each ingredient to each reaction tube, plus electrophoresis using an extended gel, which required 3 hr 30 min for running [6]. Even the cost of our modified MLVF was much lower than that of PFGE ($10 vs. $15 per test).
As shown by Simpson's diversity index, in our study, MLVF showed discriminatory power similar to that of PFGE. This finding is consistent with the findings of a previous study that used 8 different VNTR loci, including spa and sspa [14]. However, the congruence between types defined by PFGE and MLVF was fairly low, as reflected by adjusted Rand index and Wallace coefficients. The adjusted Rand index allows a quantitative evaluation of the global congruence between the 2 partitions [11]. On the other hand, Wallace coefficients, given a particular typing method, provide an estimate of how much new information is obtained from another method. The adjusted Rand index between PFGE and MLVF calculated in our study was similar to that calculated in 2 previous studies [16, 17]. However, the Wallace coefficient of PFGE predicting MLVF type in our study was lower than that calculated in the 2 previous studies. The discrepancy in the results may be explained by the difference in the S. aureus isolates used. Since the electrophoretic pattern of MLVF is based on staphylococcal interspersed repeats, the difference in the bacterial isolates might have affected the clustering profile. Those previous studies enrolled EMRSA-15 (ST22) and EMRSA-16 (ST36/ST30) or ST398 MRSA. In our study, we used various clinical isolates with diverse PFGE or MLVF patterns that could be classified into 37 types by PFGE (80% cutoff) and into 28 types, including unique types by MLVF (75% cutoff). Moreover, 15 of 65 isolates were methicillin-susceptible S. aureus (MSSA), which is known to be more polyclonal than MRSA [18].
Another explanation for the relatively low Wallace coefficients of PFGE predicting MLVF types in our study could be that we used only SmaI for DNA digestion in the PFGE. In the study by Rasschaert et al., BstZI, SacII, ApaI, and combinations of these enzymes were used [17]. The combination of all 3 restriction enzymes yielded a Wallace coefficient of 1.00 in predicting MLVF types in that study.
However, when the relationship between 2 specific isolates was analyzed, PFGE (cutoff 80%) and MLVF (cutoff 75%) showed 91.0% concordance, calculated by each cell (Table 4). In other words, every pair of isolates can be distinguished by either MLVF or PFGE. Although the InfoQuestFP software provided excellent dendrograms by applying multilinkage where branches degenerate, every dendrogram would confront loss of information while converting the data set of similarity values into a simplified hierarchical structure [19]. Furthermore, similar to PFGE analysis, MLVF also required careful visual inspection to confirm the results owing to small band shifts.
Even though the Wallace coefficients for MLVF predicting PFGE type (gold standard cutoff of 80%) were similar at both MLVF cutoffs of 75% and 70% (0.233 vs. 0.232), we suggest that a 75% cutoff be used for MLVF, since it showed higher discriminatory power.
In conclusion, our modified protocol of MLVF demonstrated good performance in typing of clinically significant S. aureus isolates selected on the basis of the CDC/NHSN criteria. Furthermore, we were able to shorten the running time by using a PCR premixture kit and performing electrophoresis using a 12-cm length system. Because MLVF has large advantage over PFGE in terms of technical aspects, MLVF could be used in place of PFGE, especially when urgent investigation of a hospital S. aureus outbreak is required.





 XML Download
XML Download