

 PDF
PDF ePub
ePub Citation
Citation Print
Print



Introduction
Regulation of trophoblast differentiation is critical for maintaining the pregnancy. Abnormalities in placental formation due to shallow trophoblast invasion may underlie the etiology of certain complications associated with human pregnancy such as preeclampsia and fetal growth restriction [1]. Trophoblast invasion is accompanied by a reduction in proliferative capacity of the cells and requires the regulated expression of either proteinases or adhesion molecules [2]. Especially, α1 integrin promotes invasion in vitro, whereas α5 integrin inhibits this process [3].
For the majority of the first trimester, the placenta develops in a hypoxic environment [4]. This hypoxic environment is important for the control of extravillous trophoblast (EVT) outgrowth as it maintains trophoblasts in a proliferative, noninvasive, and immature phenotype [2], and reduces the invasive capacity of trophoblasts and the expression of molecules associated with an invasive trophoblast phenotype such as α1 integrin [56]. Intervillous blood flow increases at around 10–12 weeks of gestation, resulting in exposure of trophoblasts to increased oxygen tension [7]. This increased oxygen tension promotes trophoblast differentiation toward the invasive pathway [28].
In recent years, hypoxia-inducible factor (HIF) has been identified as a transcription factor that controls certain target genes under low oxygen tensions [91011]. By regulating target genes, this transcription factor plays a role in angiogenesis, erythropoiesis, glycolysis, vascular tone, pH homeostasis, and cell survival decisions like proliferation and apoptosis [71012]. HIF is a heterodimer composed of an oxygen-regulated α-subunit (HIF-1α or HIF-2α) and a β-subunit (HIF-β). In the presence of oxygen, 2 prolines of the HIF-1α chain are hydroxylated by prolyl hydroxylases with oxygen acting as the substrate. The hydroxylated prolyl residues are recognized by the von Hippel-Lindau protein as a component of the E3 ubiquitin ligase, which targets HIF-1α for proteasomal degradation. In contrast, under low oxygen tensions, HIF accumulates in the cell and induces transcription of target genes [1011].
HIF-1α is known to inhibit trophoblast differentiation toward invasive EVTs [7]. HIF-1α is highly expressed in the placenta from early gestation (between 5–8 weeks) and then levels of this protein fall suddenly around gestational week 10–12 when intervillous blood flow and placental oxygenation begin to increase [13]. This decrease in the level of HIF-1α allows trophoblast differentiation into invasive EVTs that invade deep into the maternal uterus [7]. Abnormally elevated HIF-1α could result in the arrest of trophoblasts in a relatively immature state of differentiation, resulting in shallow trophoblast invasion into the uterus and reduced uteroplacental perfusion [7].
Estradiol is essential for the initiation and maintenance of pregnancy in humans as well as in experimental animals [14]. Maternal serum estradiol level begins to rise during weeks 6–8 of gestation when placental function becomes apparent. Estradiol can induce trophoblast differentiation [1516]. Recent studies reported that estradiol downregulates the expression of HIF-1α both in vivo [17] and in vitro [18].
Taken together, these results led us to hypothesize that estradiol controls trophoblast differentiation by inhibiting HIF-1α. The purpose of this study was to investigate the effects of estradiol on the expression of HIF-1α and trophoblast differentiation in human first trimester placentas.
Materials and methods
1. Explant cultures
Villous explant cultures were established from first trimester human placentas of normal pregnancy (6–8 weeks of gestation, n=3). Gestational age was calculated from the date of the last menstrual period and, if necessary, was adjusted according to ultrasonic measurements of the gestational sac and fetal crown-rump length.
All subjects provided informed consent prior to participation in the study, and this study was approved by the Institutional Review Board of Korea University College of Medicine.
All villi were from concordant pregnancies. Small pieces of tissue (2–3 mm) from the periphery of the placental debris were dissected (usually 4–8 villus tips per sample). To prepare rat-tail collagen I-coated plates (BD Biosciences, Bedford, UK), 0.8 mL of collagen I (Invitrogen, Grand Island, NY, USA) was mixed with 0.1 mL of 10×DMEM (1:10; Invitrogen) and 0.1 mL of 7.5% NaHCO4 (1:10). For each explant, 80 µL of collagen was placed in the center of a 12-well culture dish. Gels were prepared by placing a drop of the appropriate substrate in 12-well culture dishes. Matrigel (BD Biosciences) was aliquoted at 4°C and warmed to 37°C to gel. Gels were produced at least 24 hours before use and incubated in medium at 37°C to allow full contracture. After formation of the gels (30 minutes at 37°C), the dissected tissue pieces were carefully placed on top of each gel drop, were covered with 20 µL of medium, and incubated for 2–4 hours to allow for proper anchorage. Subsequently, explants were flooded with 1 mL of DMEM/F12 (Invitrogen) supplemented with 10% FBS (JR Scientific, Woodland, CA, USA) and incubated overnight at 37°C with 5% CO2. Normal range of estradiol level is 0.55–5.10 ng/mL in 6–7 weeks of gestation, respectively [19]. We adapted 1 ng/mL as physiologic level in this study. After an initial period of 24 hours, during which cytotrophoblast proliferation occurred at the villous tips, 1 ng/mL of estradiol (1,3,5[10]-Estratriene-3,17-diol, cell-culture tested; Sigma, St. Louis, MO, USA), 0.1 or 10 ng/mL of estradiol were added to the culture medium, and the explant cultures were then incubated for approximately 5 days. Control experiments were run in parallel using explants from the same placenta cultured in medium alone. Culture media were changed every 48 hours and collected. The viabilities of the explant cultures were assessed by measuring human chorionic gonadotropin levels.
2. Assessment of trophoblast outgrowth and invasion
EVT outgrowth occurred largely across the surface of the gel. Outgrowth and invasion were examined qualitatively taking into account the morphology of the constituent cells. Images were taken at different time points with a digital inverted microscope (Olympus IX71 and CKX41 camera; Olympus, Tokyo, Japan).
3. Western blotting
Protein lysates were obtained using a buffer containing 50 mM HEPES (pH 7.5), 150 mM NaCl, 1.5 mM MgCl2, 1 mM EDTA, 10% glycerol, 1% Triton X-100, and a mixture of protease inhibitors (aprotinin, phenylmethylsulfonyl fluoride, and sodium orthovanadate). Total tissues lysates were prepared by homogenization. The extracted protein concentration was measured according to the method of Bradford. Equal amounts of total protein (10 µg/sample) were resolved on a 12% sodium dodecyl sulfate-polyacrylamide gel (Bio-Rad Laboratories, Hercules, CA, USA). Proteins were transferred to a nitrocellulose membrane (HybondTM-P; Amersham Biosciences, Piscataway, NJ, USA). After blocking with TBS and 0.1% Tween-20 at 4°C overnight, the membranes were incubated with primary antibodies for anti-mouse HIF-1α (dilution 1:1,000; Santa Cruz Biotechnology Inc., Santa Cruz, CA, USA) and anti-mouse glyceraldehyde 3-phosphate dehydrogenase (dilution 1:2,000; Bio-Rad Hercules) for 24 hours followed by incubation with secondary antibodies linked to horseradish peroxidase. Immunoreactive proteins were visualized by chemiluminescence using SuperSignal West Dura Extended Duration Substrate (Pierce Chemical Co., Rockford, IL, USA), and the signals were detected on X-ray film (Agfa-Gevaert, Mortsel, Belgium).
Signals in the linear range of the film were digitized, and densitometry was performed using IMT-i-Delta (Image and Microscope Technology, Daejeon, Korea).
4. Immunohistochemical staining for integrins
To confirm the differentiation of the trophoblasts into invasive EVTs, immunohistochemistry staining for α1 and α5 integrins was performed. Briefly, tissue sections (5 µm) were deparaffinized and then rehydrated and blocked with 3% H2O2 in methanol for 30 minutes followed by universal blocking (normal serum, 1.5%; Vector Laboratories, Burlingame, CA, USA). Antibodies reactive to mouse anti-human integrin α1 (Santa Cruz Biotechnology Inc.) or rabbit anti-human integrin α5 (Santa Cruz Biotechnology Inc.) were used at dilutions of 1:100 and were incubated with the samples for 1 hour. After the primary antibodies were applied, the slides were incubated overnight at room temperature. Detection was performed using a secondary antibody (Vector Laboratories). All samples were counterstained with Mayer's hematoxylin before mounting with Immunomount (Lab Vision, Fremont, CA, USA).
Results
1. Effects of estradiol on outgrowth and invasion of EVTs
Villous explants from gestational weeks 6 to 8 were maintained in culture for 5 days under 3% O2 tension with 1 ng/mL of estradiol. Control experiments were run in parallel using explants from the same placenta cultured in medium alone. EVTs emerging from first trimester villous explant cultures formed outgrowths of cells from the distal end and invaded the surrounding Matrigel (Fig. 1). Exposure of villous explants to estradiol increased EVT invasion into the surrounding Matrigel (arrow, Fig. 1D) when compared to control villous explants, which showed the typical low O2 tension-induced outgrowth (arrowhead, Fig. 1C).
 | Fig. 1Effects of estradiol on extravillous trophoblast outgrowth and invasion in first trimester villous explant cultures. Dissected villous tissues were cultivated on Matrigel under 3% O2 tension (A, C) or under 3% O2 tension with 1 ng/mL of estradiol (B, D). A series of images were taken under an inverted microscope at ×10 magnification at day 0 (A, B) and day 5 (C, D).
|
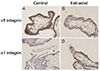
2. Effects of estradiol on EVT differentiation
To confirm the differentiation of trophoblasts into invasive EVTs, we performed immunohistochemical staining for α1 and α5 integrins (Fig. 2). A few cells immunopositive for α5 integrin were noted in explants cultured with 1 ng/mL of estradiol under 3% O2 tension. However, strong positive immunoreactivity for α1 integrin was observed in explants cultured with 1 ng/mL of estradiol when compared to control villous explants.
3. Effects of estradiol on the expression of HIF-1α in villous explant cultures
To determine whether estradiol plays a role in modulating the expression of HIF-1α, we studied the effect of estradiol on the expression of HIF-1α in villous explant cultures (Fig. 3). On western blots, the protein expression of HIF-1α was detected in both untreated villous explants under 3% O2 tension and with 1 ng/mL of estradiol (physiologic level) under 3% O2 tension. The expression of HIF-1α in villous explant cultures from gestational weeks 6 to 8 decreased significantly after treatment with 1 ng/mL of estradiol under 3% O2 tension when compared to the control villous explants (Fig. 3A). Corresponding densitometric analysis confirmed these results (Fig. 3B).
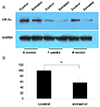 | Fig. 3Western blot analysis of hypoxia-inducible factor (HIF)-1α in villous explant cultures. (A) The expression of HIF-1α from gestational weeks 6 to 8 villous explants were cultured under 3% O2 tension with 1 ng/mL of estradiol for 5 days and were compared with that of control explant cultures. (B) Densitometric analysis. Data are expressed as a percentage of control values (explants cultured under 3% O2 tension) of 3 separate experiments in triplicate.
GAPDH, glyceraldehyde 3-phosphate dehydrogenase.
a)P<0.05 vs. control.
|
4. Effects of different doses of estradiol on the expression of HIF-1α
Next, to investigate the expression of HIF-1α at different doses of estradiol, villous explants were cultured with 0.1 or 10 ng/mL of estradiol under 3% O2 tension for 5 days, and the expression of HIF-1α was compared with that of control explant cultures (Fig. 4). The expression of HIF-1α from villous explants cultured with 1 ng/mL of estradiol was lower than that of untreated villous explants. However, the expression of HIF-1α from explant cultures treated with 0.1 or 10 ng/mL of estradiol was greater than that of control villous explants (Fig. 4A). Corresponding densitometric analysis confirmed these results (Fig. 4B).
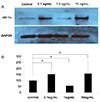 | Fig. 4Western blot analysis of hypoxia-inducible factor (HIF)-1α in villous explant cultures. (A) Expression of HIF-1α from villous explants cultured under 3% O2 tension with 0.1–10 ng/mL of estradiol for 5 days was compared with that of control explant cultures. (B) Densitometric analysis. Data are expressed as a percentage of control values (explants cultured under 3% O2 tension) of 3 separate experiments performed in triplicate.
GAPDH, glyceraldehyde 3-phosphate dehydrogenase.
a)P<0.05 vs. control.
|
Discussion
To the best of our knowledge, this study was the first to report that a physiologic level of estradiol downregulates the hypoxic induction of HIF-1α, and promotes trophoblast differentiation toward invasive EVTs in first trimester villous explant cultures. The HIF-1α protein is significantly overexpressed in preeclamptic placentas [20], and villous explants from preeclamptic placentas fail to adequately downregulate HIF-1α protein expression upon oxygenation [21], although the cause of aberrant HIF-1α expression in preeclamptic placentae remains to be elucidated. Estrogen is essential for the initiation and maintenance of pregnancy [14]. Especially, it has been known that estrogen influences placental growth and increases the fetoplacental blood flow [2223]. The association of estradiol with preeclampsia has been also suggested from several studies, although the mechanism underlying the association is still unclear. Several studies measuring estradiol in blood from preeclamptic women have reported lower estradiol levels [242526] than in the controls, while others have not found a difference [272829]. Moreover, some studies have reported a positive effect of estradiol administration on severe preeclampsia [3031]. Reduced estrogen exposure has been hypothesized to explain the lower risk of breast cancer found in women with preeclamptic pregnancies [32] and in their daughters [33].
Our study suggests that inappropriate levels of estradiol could contribute to an elevation in the placental levels of HIF-1α, which could lead to the shadow invasion of trophoblasts into the spiral arteries and uterine wall, as seen in preeclampsia. Hypoxia and placental insufficiency may lead to a deficiency in placental estrogen, which in turn may result in further elevation of placental HIF-1α levels [2734], resulting in a vicious cycle. Elevated levels of HIF-1α induced by inappropriate levels of estradiol may induce the transcription of target genes, such as soluble vascular endothelial growth factor receptor-1, which plays a major role in the pathogenesis of preeclampsia [35].
The molecular mechanisms by which estradiol regulates expression of HIF-1α in explant cultures have yet to be determined, but we propose the following hypotheses. First, the effects of estradiol on HIF-1α may be due to activation of one or both of the 2 estrogen receptors (ERs), ERα and ERβ. Estradiol attenuated the hypoxic induction of HIF-1α in Hep3B cells, but an ER antagonist eliminated these effects of estradiol, indicating that estradiol may downregulate HIF-1α through an ER-dependent mechanism [18]. It is known that homozygous T-A haplotype carriers of ERα PvuII and XbaI gene polymorphisms have an increased risk of severe preeclampsia [36], and a single nucleotide polymorphism of the ERβ gene was linked to preeclampsia after stratification according to the presence of a family history of hypertension [37]. These may be possible reasons why studies measuring the levels of estradiol in the blood of preeclamptic women have reported conflicting results.Second, estradiol may control HIF-1α via 2-methoxyestradiol (2-ME). 2-ME is a natural metabolite of estradiol and possesses antitumor and antiangiogenic activity [38]. 2-ME has low affinities for ERα and ERβ, and its antiproliferative activity is independent of any interaction with these receptors [39]. 2-ME destabilizes microtubules and downregulates HIF-1α [40]. During pregnancy, 2-ME is generated by catechol-O-methyltransferase (COMT) in the placenta [41], and its maternal serum level increases during the third trimester of normal human pregnancy [42]. However, the placental activity of COMT is suppressed in women with preeclampsia [43], and the circulating level of 2-ME is lower in women with preeclampsia than in normal pregnant women [44]. It has been reported that pregnant mice deficient in COMT developed multiple functional and structural features consistent with a preeclampsia-like phenotype due to the absence of 2-ME [44]. Therefore, the estradiol-mediated attenuation of HIF-1α may be a function of the production of 2-ME by COMT in villous explant cultures. However, we did not evaluate ER status nor the expression levels of 2-ME or COMT in villous explant cultures. Therefore, further studies are required to better understand the molecular mechanisms by which estradiol regulates the expression of HIF-1α in pregnancy.
In this study, a physiologic level of estradiol (1 ng/mL) decreased the expression of HIF-1α, whereas 0.1 and 10 ng/mL of estradiol increased the expression of HIF-1α when compared to control villous explants. The mechanism underlying these results is unclear. Some studies have reported that estradiol downregulates the expression of HIF-1α [1718]. Conversely, others have reported that estradiol increases the expression of HIF-1α [45]. It seems likely that the effects of estradiol on HIF-1α may differ according to the concentration of estradiol employed.
HIF-1α is highly expressed in the placenta and maternal serum estradiol level begins to rise at around 5–8 weeks of gestation. However, as intervillous blood flow increases at around 10–12 weeks of gestation, levels of HIF-1α fall suddenly and trophoblast was differentiated toward invasive pathway [13]. This study was to investigate the effects of estradiol on expression of HIF-1α and differentiation of trophoblast. Therefore, to control these characteristics of placentas acquired at around 10–12 weeks of gestation, we used the placentas from 6 to 8 weeks gestations.
In conclusion, we suggest that physiologic levels of estradiol may downregulate expression of HIF-1α, which in turn promote trophoblast differentiation into invasive phenotype. This raises the possibility that inappropriate levels of estradiol might contribute to major complications of pregnancy that are associated with abnormal trophoblast invasion and placental development, such as preeclampsia. Further studies are needed to evaluate the role of estradiol on pathophysiology of preeclampsia and clinical effects of administration with estradiol on preeclamptic patients.
 XML Download
XML Download