

 PDF
PDF ePub
ePub Citation
Citation Print
Print



Introduction
Premature birth occurs in 7%–10% of all pregnancies and is a major cause of perinatal morbidity and mortality [1]. In order to prevent and control premature birth, a more precise understanding of the mechanisms underlying parturition is required. The uterine myometrium undergoes a large number of physiological and molecular changes during pregnancy. It maintains a relatively quiescent state for most of the pregnancy, but from the late third trimester, as the uterine muscles starts getting activated in order to prepare for labor, the uterine myometrium facilitates birth through strong, synchronous contractions accompanying the start of labor.
During pregnancy, potassium ion channels play an important role in the activation of uterine muscles [23]. The outward flow of potassium ions caused by potassium ion channel opening repolarizes the cell membrane potential, reducing intracellular the influx of calcium ions through the cell membrane of sarcolemmal voltage-dependent calcium channels, thereby reducing cellular excitability and contractility [2]. Therefore, changes in potassium ion channel expression or activity can lead to changes in myometrial excitability and contractility. Studies have reported that potassium channel-opening drugs impair human uterine smooth muscle contraction; these drugs have been suggested as a new class of tocolytics [456]. The myometrium is reported to contain various potassium ion channels, including large-conductance calcium- and voltage-sensitive K+ channels, adenosine triphosphate (ATP)-sensitive K+ channels, shaker-like voltage-gated potassium channels, and small-conductance calcium-sensitive potassium channels [3]. Animal and human in vitro studies have shown that, of these potassium channels, ATP-sensitive potassium (KATP) channels are closely related to myometrial activation [3789]. KATP channels are heteromultimers consisting of inwardly rectifying K+ channel (Kir) subunits and regulatory sulfonylurea receptor (SUR) subunits. Different combinations of 3 types of SUR subunits (SUR1, SUR2A, and SUR2B) and 2 types of Kir subunits (Kir6.1 and Kir6.2) result in the expression of different subtypes in various tissues, including human smooth muscle [10]. The majority of KATP channels in the human uterine myometrium are composed of SUR2B and Kir6.1 [1112].
Corticotropin-releasing hormone (CRH) is a 41-amino-acid neuropeptide that is produced in the placenta and fetal membranes during human pregnancy [13]. The concentration of CRH in the maternal plasma begins to increase rapidly during the third trimester of pregnancy, reaching a peak during labor [14151617]. An earlier and more rapid increase in plasma CRH levels has been reported in premature birth, and a slower increase has been reported in post-term pregnancy [17181920]. This indicates that increase in plasma CRH concentration is closely related to the onset timing of parturition, and therefore CRH has been proposed to regulate the placental clock.
Since CRH is related to the onset timing of parturition, we predicted that it would affect expression of KATP channels, which are themselves related to myometrial activation. To our knowledge, this study is the first to investigate CRH concentration-dependent changes in the expression of the KATP channel subunits Kir6.1 and SUR2B in human term non-laboring myometrium.
Materials and methods
1. Uterine myometrial smooth muscle cell isolation and cultures
Samples for the study were obtained from term pregnant women (37–42 weeks; average age, 32 years; n=5) who had no underlying disease (e.g., hypertension, diabetes, preeclampsia, intrauterine growth restriction) and were undergoing elective cesarean section. The study was approved by the Research Ethics Committee of Eulji University Hospital in Korea. Informed consent for the sampling of uterine tissue was obtained from each patient. We used scissors or a knife to obtain uterine tissue from the patients; a 1×1×1-cm piece of tissue was removed from the upper lip of the uterine incision. After immediately removing the serosa and endometrium with fine scissors, the tissue collected was washed in Hanks' balanced salt solution (GIBCO BRL, Gaithersburg, MD, USA) before cutting it into 3×3-mm samples. These tissue samples were cultured for 16 hours at 37°C in DMEM (GIBCO BRL) containing 1 mg/mL collagenase II (GIBCO BRL) and 1 mg/mL deoxyribonuclease I (Sigma-Aldrich, St. Louis, MO, USA). After the nondispersed tissue fragments had been dispersed in the culture medium using a pipette, the medium was filtered using 4 sheets of sterilized gauze. The filtered medium was subjected to centrifugation at 600 ×g for 10 minutes. The cell pellets were placed in a 100-mm dish containing DMEM with 10% fetal bovine serum and 1% penicillin-streptomycin, before culture at 37°C and 5% CO2, in a 95% humidified atmosphere. Flow cytometry was performed using smooth muscle actin B-4 (SC-53142; Santa Cruz Biotechnology, Santa Cruz, CA, USA), a smooth muscle-specific antibody, in order to check the purity of smooth muscle cells; average purity of over 99% was confirmed (data not shown).
2. CRH treatment
The subcultured human myometrial smooth muscle cells (HSMCs) were shifted again to 6-well dishes, and after culture to at least 50% confluency, they were treated with CRH (Sigma-Aldrich) for 24 hours. When cells were treated with CRH for 6, 12, 24, and 48 hours, the results for 12 hours and more were similar and therefore a consistent treatment duration of 24 hours was chosen for convenience. To determine the changes in blood CRH concentration during pregnancy, we treated cells with 6 different concentrations; the final concentrations of CRH in each well were 0, 1, 10, 102, 103, and 104 pmol/L. The cells were pretreated with a CRH receptor-1 antagonist (antalarmin) and a CRH receptor-2 antagonist (astressin 2b) (Sigma-Aldrich) for 1 hour, at a concentration of 10−7 mol/L, before CRH treatment.
3. Total RNA extraction and reverse transcription (RT)-polymerase chain reaction (PCR)
After HSMCs were treated with different concentrations of CRH for 24 hours, an RNeasy Plus Mini Kit (Qiagen, Valencia, CA, USA) was used according to the manufacturer's instructions to isolate total RNA. Total RNA concentrations were measured spectrophotometrically at optical densities of 260 and 280 nm. After quantification, 2 µg of RNA was reverse transcribed with oligo dT random primers in a final volume of 25 μL using murine leukemia virus reverse transcriptase (Bioneer Corp., Daejeon, Korea) and the resultant cDNA was stored at −20°C. The cDNA was amplified by PCR, which was performed in a Gene Amp PCR system 9600-R (Applied Biosystems, Carlsbad, CA, USA). The specific primers used for the amplification were as follows [21]: Kir6.1: sense, 5′-CCTGAAGTGGCGTCACAC-3′ and antisense, 5′-GTCACCCACCCGGAACATGA-3′; SUR2B: sense, 5′-TCCGCTCTAGACTGTCCATCATTC-3′ and antisense, 5′-TCCTCTCTTCATCACAATGACCAGGTC-3′; and β-actin: sense, 5′-CCAGACAGCACTGTGTTG-3′ and antisense, 5′-CTGAGAGGGAAATCGTGC-3′. The primers were synthesized by Bioneer Corp. The reaction solution consisted of 2.0 μL of diluted cDNA, 0.2 μM each paired primer, and 1X Taq PCR MasterMix (Qiagen). The annealing temperature was set at 57°C, 61°C, and 55°C for Kir6.1, SUR2B, and β-actin, respectively; 32, 36, and 22 cycles, respectively, were used for these PCRs. All the amplified products were separated on 2.0% agarose gels and bands were visualized with ethidium bromide staining. RT-PCR was used to evaluate mRNA expression for the KATP channel subunits Kir6.1 and SUR2B according to CRH concentrations. This experiment was performed 5 times, using HSMCs from different patients.
4. Protein isolation and western blotting analysis of Kir6.1 and SUR2B protein expression
CRH-treated cells in 6-well dishes were washed twice with cold phosphate-buffered saline and then lysed in cold lysis buffer Proprep (iNtRON, Seongnam, Korea) for 30 minutes in ice. The cell lysates were centrifuged at 13,000 ×g for 30 minutes at 4°C. The supernatant was collected and the protein concentration was assayed using a bicinchoninic acid assay. Samples were then aliquoted and stored at −80°C. Electrophoresis was carried out using 60 μg aliquots of protein samples that were denatured by adding LDS Sample Buffer and Sample Reducing Agent (Invitrogen, Carlsbad, CA, USA) and heating for 10 minutes at 70°C. Aliquots of samples were separated on a Bolt 4%–12% Bis-Tris Plus gel (Invitrogen), and the proteins were electrophoretically transferred to a nitrocellulose membrane with iBlot Dry Blotting System (Invitrogen). The membrane was then blocked using Tris-buffered saline containing 0.1% Tween20 and 5% nonfat milk powder (wt/vol) for 1 hour at room temperature. After 3 washes with Tris-buffered saline containing 0.1% Tween 20, the membrane was incubated with goat anti-human Kir6.1 (1:200; Santa Cruz Biotechnology), goat anti-human SUR2B (1:200; Santa Cruz Biotechnology), and goat anti-human β-actin (1:10,000; Santa Cruz Biotechnology) antibodies at 4°C overnight. The membrane was then incubated with a horseradish peroxidase-conjugated rabbit anti-goat IgG (1:5,000) for 1 hour at room temperature after 3 washes. Immunoreactive proteins were visualized with the enhanced chemiluminescence western blotting detection system ImageQuant LAS 4000 mini (GE Healthcare Bio-Sciences, Uppsala, Sweden). Western blotting was performed 5 times with different HSMCs.
5. Measurement of intracellular Ca2+
HSMCs were preloaded with 10 μM Fluo-8 AM (AAT Bioquest, Sunnyvale, CA, USA) in DMEM (GIBCO BRL) at 37°C for 60 minutes. Preloaded cells were washed twice with Hanks and HEPES buffer (GIBCO BRL) containing probenecid (2.5 mM) to remove excess probes. Cells were first treated with 1 or 103 pmol/L CRH for 24 hours, after which changes in cytosolic Ca2+ levels over time were assessed using a fluorescence plate reader (Infinite M1000; Tecan, Mannedorf, Switzerland) with increasing concentrations of oxytocin. The final concentrations of oxytocin were 10−9, 10−8, 10−7, and 10−6 M. Fluorescence intensity was measured for 2,000 seconds with excitation at 490 nm and emission at 525 nm. This experiment was performed 3 times with different myometrial smooth muscle cells.
Results
1. Effects of CRH on the expression of Kir6.1/SUR2B transcripts at different concentrations of CRH
The mRNA expression of Kir6.1 and SUR2B at 1 pmol/L CRH was significantly higher than that for the control (P<0.05). mRNA expression decreased at concentrations of 10 and 102 pmol/L CRH and a level similar to that in controls was noted; then, at 103 and 104 pmol/L, the expression decreased even further, and this decrease was statistically significant compared to the levels seen in controls (Fig. 1A). Protein expression of Kir6.1/SUR2B showed a similar pattern to mRNA expression, but unlike mRNA levels, protein levels did not decrease significantly compared to those of the controls at 103 and 104 pmol/L (Fig. 1B).
Fig. 1
Molecular expression of Kir6.1 and SUR2B with 1–104 pmol/L corticotropin-releasing hormone (CRH) in human myometrial smooth muscle cells. Blots are representative of 5 experiments. The expression level is presented as the ratio of the radioactivity of the band to that of the corresponding β-actin band. Values are presented as mean±standard error for 5 cultures from 5 patients. (A) mRNA expression of Kir6.1 and SUR2B was measured by reverse transcription-polymerase chain reaction. (B) Protein expression of Kir6.1 and SUR2B was measured by western blot analysis.
a)P<0.05 compared with control.
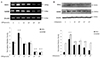
2. Pretreatment effects of CRH receptor-1/receptor-2 antagonists on the expression of Kir6.1/SUR2B mRNA at low and high CRH concentrations
At a CRH concentration of 1 pmol/L, the increase in Kir6.1 mRNA showed no difference when the cells had been pretreated with antalarmin or astressin 2b, but the increase in SUR2B mRNA expression was reduced following pretreatment with either antalarmin or astressin 2b (Fig. 2A). In particular, the increase in SUR2B mRNA was lower following antalarmin pretreatment compared to the results seen for astressin 2b treatment (P<0.05). At a CRH concentration of 104 pmol/L, the decrease in Kir6.1 mRNA was unaffected by antalarmin or astressin 2b pretreatment, but the decreased SUR2B mRNA expression was negated by pretreatment with either antalarmin or astressin 2b (Fig. 2B). In particular, the effect of astressin 2b pretreatment was significantly greater than that of antalarmin pretreatment (P<0.05).
Fig. 2
Effects of corticotropin-releasing hormone (CRH)-R1/-R2 antagonists on mRNA expression of Kir6.1 and SUR2B at low (1 pmol/L) (A) and high (104 pmol/L) (B) concentrations of CRH. mRNA expression of Kir6.1 and SUR2B was measured by reverse transcription-polymerase chain reaction. Cells were pretreated with CRH-R1 antagonist (antalarmin) and CRH-R2 antagonist (astressin 2B) for 1 hour before treatment with CRH. Thereafter, cells were harvested for mRNA analysis. Representative bands are presented. The expression level is presented as the ratio of the radioactivity of the band to that of the corresponding β-actin band. Values are presented as mean±standard error for 5 cultures from 5 patients.
a)P<0.05.

3. Effects of different concentrations of CRH on CRH-dependent changes in intracellular Ca2+ concentration following oxytocin treatment
When HSMCs are treated with oxytocin, the intracellular Ca2+ concentration increased with increase in oxytocin concentration. The intracellular Ca2+ in HSMCs treated with 1 pmol/L CRH for 24 hours decreased significantly compared to that in the control (P<0.05) (Fig. 3). At a CRH concentration of 103 pmol/L, the Ca2+ concentration did not significantly differ from that for the control.
Fig. 3
Effects of corticotropin-releasing hormone (CRH) on oxytocin-induced increases in intracellular Ca2+ concentrations in human myometrial smooth muscle cells (n=3). CRH treatment was performed 24 hours before treating cells with oxytocin (OXY) and measuring intracellular Ca2+ using Fluo-8 AM ester. The Ca2+ traces were obtained by connecting the experimental value measured every 5 seconds. The representative traces are shown. Each value represents the mean measured for 3 replicate wells from one 96-well plate.
a)P<0.05.

Discussion
There is a considerable change in the maternal plasma CRH concentrations throughout pregnancy. Campbell et al. [22] reported that CRH concentrations begin to increase from 26 to 28 weeks (50±15 pg/mL), reach 1,462±182 pg/mL at term, and peak at 1,680±101 (±standard error of the mean) pg/mL during active labor. Sasaki et al. [15] reported that the CRH concentration in the first trimester was similar to that for non-pregnant women, at 5.8±0.8 pg/mL (range, 3.3–10.8), before increasing to 800±163 pg/mL in the late third trimester, and to 4,409±591 pg/mL (range, 1,231–13,910) during active labor. The CRH concentrations used in the present study were based on the results of Sasaki et al. [15], with the lowest concentration of 1 pmol/L corresponding to the first trimester (5.8 pg/mL=1.24 pmol/L), and 103 pmol/L corresponding to the peak maternal plasma concentration (4,409 pg/mL=926.75 pmol/L) during labor.
The hormone CRH controls the physiological events leading to parturition; studies have reported that it is likely to control myometrial contractility in relation to the timing of birth [232425262728]. CRH is reported to have a dual action, where it promotes myometrial quiescence for most of the pregnancy, but facilitates myometrial contractility after the onset of parturition [23272829]. CRH also increases the contractile effects of oxytocin in pregnant myometrium, but there are conflicting reports indicating that it can show relaxant effects [3031]. In an in vitro study, Zhang et al. [24] reported that CRH did not show a relaxant effect in the laboring myometrium, but did have a relaxant effect on spontaneous contractions in the non-laboring term myometrium, which was mediated by CRH receptor-1. Recently, studies have been performed to determine the mechanisms underlying CRH dual action during pregnancy. Hillhouse and Grammatopoulos [27] reported that, after binding to the receptor, CRH could activate multiple classes of G proteins, which could lead to multiple signaling pathways, and that contrasting effects could be achieved during the course of pregnancy by changes in the pattern of activated G proteins by post-receptor modifications. Thus, the dual action of CRH in the myometrium has various explanations in terms of binding to CRH-R1 or CRH-R2, or in terms of activated G-protein classes, but the definite mechanism still needs to be determined.
In our study, dual action of CRH was observed, with Kir6.1 and SUR2B mRNA expression increasing significantly compared to that of the control at a low CRH concentration of 1 pmol/L, and decreasing significantly at a high CRH concentration of over 103 pmol/L (Fig. 2). Thus, at low concentrations corresponding to maternal plasma CRH levels in early pregnancy, KATP channel expression increased significantly, acting to stabilize the myometrial membrane resting potential and inhibiting myometrial activation. We confirmed the possibility that, from the late third trimester, when plasma CRH levels increase, expression of KATP channels begins to reduce, causing myometrial activation and contributing to contractions at the onset of labor, when expression is at its lowest. Although there have previously been various studies related to the dual action of CRH during pregnancy, our study showed that a difference in CRH concentration alone can produce opposite actions, increasing or decreasing KATP channel expression, and that this could be another explanation for the dual action of CRH activating or relaxing uterine smooth muscles during pregnancy. In an experiment to determine which CRH receptor was involved in the reaction, we found that Kir6.1 mRNA expression was unaffected by pretreatment with antalarmin or astressin 2b at both high and low concentrations of CRH. However, SUR2B mRNA expression was affected, with the increase in expression at low CRH concentrations antagonized by pretreatment with antalarmin, and the decrease in SUR2B mRNA expression at high CRH concentrations antagonized by pretreatment with astressin 2b. CRH has a higher affinity for CRH receptor-R1 than for CRH receptor-R2 [323334]. Some studies showed that CRH receptor-R1 activation could promote myometrium quiescence, whereas CRH receptor-R2 activation could promote myometrial contractility [242835]. Hence, we surmise that, at low concentrations CRH binds preferentially to CRH receptor-1, causing an increase in expression, and that, at high concentrations, CRH is also able to bind to CRH receptor-2, acting to reduce expression. However, the expression of Kir6.1, which is the direct channel for K+ ions, is unaffected by antalarmin or astressin 2b at high and low concentrations of CRH. In adult cardiomyocytes and rat embryonic heart H9c2 cells, upregulation of the SUR subunit alone is sufficient to increase numbers of sarcolemmal KATP channels [3637]. Hence, increased expression of SUR2B results in increased numbers of KATP channels [3839].
The results did not clearly indicate which CRH receptor and what molecular mechanisms were involved in the dual actions of CRH. Additional studies will be required to clarify the relationship between CRH receptors and KATP channel expression. Although the pattern of protein expression was similar to that of mRNA expression, there were some differences. KATP channel subunit mRNA was first cloned from the human myometrium in 2002, but protein levels could not be detected until 2011, when protein was first confirmed in the human myometrium by Xu et al [12]. The reason that KATP channels cannot be easily detected in the human myometrium at the protein level is thought to be either because they exist at a very low density, or because the mRNA is unstable and is not all translated into protein [212]. In our study as well, we were only able to detect protein expression with difficulty; we used SDS-PAGE gels with a high protein loading content of 60 µg and a high concentration of primary antibody. For the above reasons, although protein expression showed a similar pattern to mRNA expression, we think it is possible that there may be some differences, and therefore the mRNA results should be given more importance. Therefore, we have omitted the results for Kir6.1 and SUR2B protein expression after pretreatment with CRH-receptor antagonists, since they were not reliable.
In conclusion, our study demonstrates that a difference in CRH concentration alone can alter expression of KATP channels, which are related to myometrial activation, in human term myometrium that has not undergone labor. KATP channel expression shows different patterns at the low CRH concentrations of early pregnancy and the high CRH concentrations at parturition. We confirmed that a low concentration of CRH causes an increase in KATP channel expression, which may act to inhibit activation of uterine smooth muscle, while the high concentration of CRH at the start of parturition causes a decrease in KATP channel expression, which may act to increase uterine smooth muscle activation and cause contractions.
 XML Download
XML Download