

 PDF
PDF ePub
ePub Citation
Citation Print
Print



Introduction
Müllerian inhibiting substance (MIS) is known as anti-Müllerian hormone (AMH) and was named in 1946 when Alfred Jost discovered that a molecule, which was not testosterone and secreted from the rabbit embryonic testis, degenerated the Müllerian duct [1]. This hormone is a 140 kDa glycoprotein composed of 545 amino acids that belongs to the multigene family of transforming growth factor-beta (TGF-β) [2]. The Müllerian duct develops into the fallopian tube, uterus, cervix, proximal part of the vagina, and ovarian epithelium in women. MIS/AMH, which is produced at a high concentration in fetal testis, binds to the MIS/AMH receptor of the primordial Müllerian duct and suppresses development of female genitalia in boys; thus, playing an important role in sexual development [34]. In female, MIS/AMH is produced by the granulosa cells of ovary and is secreted into the follicular fluid and serum [5678]. MIS/AMH appears at the end of the third trimester of pregnancy about 36 weeks of gestation and is rarely detectable at birth and reaches the highest values after puberty [5678]. After menopause, secretion decreases, so it has also been used as a measure of the degree of loss of ovarian function [5678].
MIS/AMH not only plays an important role in fetal sexual development, but also inhibits growth of tumors originating from the Müllerian duct where the MIS/AMH receptor is expressed [91011], such as ovarian [121314151617], cervical [18192021], and endometrial cancers [2223]. Masiakos et al. [14] reported that MIS/anti-Müllerian hormone type II receptor (AMHRII) is expressed at a high frequency in cells detected in ascites of patients with stage III/IV ovarian cancer, and MIS/AMH cause the growth inhibition of ovarian cancer cells expressing the MIS/AMH receptor. Similarly, Barbie et al. [21] reported that MIS/AMHRII is expressed in a number of cervical cancer cell lines and that MIS/AMH inhibits growth of cervical cancer cells. In addition to inhibiting malignant tumors, MIS/AMH also inhibits benign diseases originating from the Müllerian duct, such as endometrial stromal cells in endometriosis [24].
Uterine leiomyoma is the most common tumor of the female reproductive tract and is a monoclonal myometrial tumor originating from the Müllerian duct [2526]. Adenomyosis is a disease in which the endometrial tissue invades the myometrium resulting in a diffusely enlarged uterus [27]. Both are benign uterine diseases of Müllerian duct origin, and they are very common in women of childbearing age. However, unlike malignant diseases, few studies have investigated MIS/AMHRII expression in myoma and adenomyosis.
In this study, we investigated MIS/AMHRII expression levels in uterine myoma and adenomyosis tissues to evaluate the possibility of using MIS/AMH as a potential therapeutic modality via receptor-mediated mechanism in uterine myoma and adenomyosis.
Materials and methods
1. Study objectives
We included 57 women who underwent myomectomy for uterine myoma (7 cases) or total hysterectomy for uterine myoma (22 cases) or adenomyosis (28 cases) from March 2016 to October 2016 at Uijeongbu St. Mary's Hospital. The patients were explained the objectives of study and informed consent was obtained from all participants. We examined 22 myometria, 29 uterine myomas, 28 endometria, and 28 adenomyosis tissue samples confirmed by a pathologist. In myomectomy, the extraction of normal muscle tissue may cause unnecessary injury to the uterus. Thus, normal muscle tissue was collected only in the hysterectomy group of myoma. This study was approved by the Institutional Review Board-Human Research Committee at Uijeongbu St. Mary's Hospital (No. UC14TISI0087).
2. Study methods
1) Reverse transcriptase-polymerase chain reaction (RT-PCR)
Normal myometrium, endometrium, uterine myoma, and adenomyosis tissues were subjected to RT-PCR to determine the Müllerian inhibiting substance type II receptor (MISRII) mRNA expression levels.
(1) RNA extraction and reverse transcription
Tissues for RNA analysis were immediately snap frozen in CryoTubes (Nunc, Roskilde, Denmark) and stored in liquid nitrogen until further processing. Total RNA was extracted from tissues using TRIZOL LS Reagent (Invitrogen, Carlsbad, CA, USA) and RT-PCR was conducted to synthesize cDNA from RNA with the cDNA Synthesis Kit (Takara Korea Biomedical Inc., Seoul, Korea). The polymerase chain reaction (PCR) mixture consisted 4 µL of 5×PCR buffer, 1 µL of Moloney murine leukemia virus reverse transcriptase, 1 µL of dNTP mix, 2 µL of random primers, 1 µL of RNase inhibitor, 2 µg of RNA, and diethyl dicarbonate water in a total volume of 20 µL. Amplification started with the DNA thermal cycler at 25°C for 10 minutes, followed by 42°C for 60 minutes, and 70°C for 10 minutes. The PCR products were stored at −20°C.
(2) RT-PCR
cDNA products were amplified using by Premix Taq (Takara Ex Taq version; Takara Korea Biomedical Inc.). The PCR mixture consisted of 10 µL of PCR premix, 1 µL of downstream primer (10 pmol), 1 µL of upstream primer (10 pmol), 1 µL of reverse transcribed cDNA, and RNase free sterile water in a total volume of 20 µL. Amplification started with denaturation at 94°C for 5 minutes followed by 30 cycles of 94°C for 40 seconds, 57°C for 40 seconds and 72°C for 25 seconds. The last reaction was performed at 72°C for 10 minutes.
The PCR primer sequences were: sense primer 5′-CCCTGCTACAGCGAAAGAAC-3′ (MISRII cDNA; Gene Bank accession no. AF172932; sequence 581–600), and anti-sense primer 5′-TGGGTCAAGTAGTGGCACAG-3′ (sequence 921–941). The PCR products (10 µL) were separated by 1.5% agarose gel electrophoresis, and the band size (361 bp) was compared with a DNA ladder.
2) Immunohistochemistry
Paraffin embedded tissue was sliced to a 4 µm thickness and attached to a ProbeOn Plus microscopic slide (Fisher Scientific, Pittsburgh, PA, USA). Tissues for immunohistochemical detection of MISRII were processed using the capillary tube method, a fast temperature-controlled machine, and the microprobe immunostaining station (Biomeda Co., Foster City, CA, USA). The immunostaining procedure was as follows: the slides were preheated at 85°C for 30 minutes, deparaffinized 3 times with xylene for 5 minutes at room temperature, and dehydrated in a graded alcohol series. The slides were autoclaved at 121°C for 10 minutes in citrate buffer for antigen retrieval (Invitrogen) and cooled for 20 minutes at room temperature. After fours washes in phosphate buffered saline (PBS), the slides were stained using the Histostain-Plus AEC kit (Invitrogen). After treatment with peroxidase to block non-specific protein binding followed by a wash in PBS, 10% goat non-immune serum was added for 60 minutes. The slides were incubated with rabbit polyclonal anti-human MISRII antiserum (provided by Dr. David T. MacLaughlin, Massachusetts General Hospital, Boston, MA, USA) at 4°C overnight. The slides were rinsed 3 times in Tris-buffered saline with 0.1% Tween 20 (T-TBS; LabVision, Fremont, CA, USA) for 5 minutes and incubated with biotinylated anti-rabbit immunoglobulin G (Invitrogen) as the secondary antibody at room temperature for 2 hours. After rinsing in T-TBS, the Streptavidin HRP detection system (Invitrogen) was applied to the slides for 30 minutes to induce the biotin-streptavidin binding reaction. After washing for 5 minutes in T-TBS, the slides were treated with 3-amino-9-ethylcarbazole for 10 minutes at room temperature, counterstained with hematoxylin, and mounted with Clearmount (Invitrogen).
3) Expression scoring system and statistical analysis
All immunostained sections were reviewed and scored independently by 2 pathologists in a blinded manner without knowledge of the clinicopathological information by considering staining intensity and the percentage of positively stained cells. The total number of cells and cells stained at each intensity were counted in each of 5 fields. Staining intensity greater than that seen in normal follicular granulosa cells was determined to be positive. The frequency of positively stained cells from 5 areas under a high magnification field were calculated as a percentage. Frequencies <30% were weakly positive, 30%–60% was moderately positive, and >60% was strongly positive. Negative was scored as 0, weakly positive as 1, moderately positive as 2, strongly positive as 3. Data were analyzed by the Kruskal-Wallis and Tukey's tests using ranks. A P<0.05 was considered statistically significant.
Results
1. Expression of MIS/AMHRII mRNA by RT-PCR
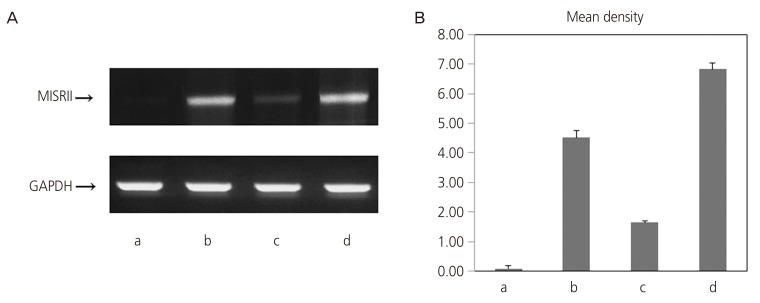
The MIS/AMHRII mRNA expression levels in uterine myometrium, endometrium, uterine myoma, and adenomyosis tissues were examined by RT-PCR. In all tissues, a band of the same 361 bp as a part (581–941) of the human MIS/AMHRII cDNA sequence (MISRII cDNA; Gene Bank Accession No. AF172932) was detected. MIS/AMHRII mRNA expression was stronger in uterine myoma than that in uterine myometrium (4.51±0.26 vs. 0.08±0.09; P<0.05). Similarly, the MIS/AMHRII mRNA expression level in adenomyosis was higher, compared with that in normal endometrial tissue (6.84±0.20 vs. 1.63±0.06; P<0.05) (Fig. 1).
Fig. 1
(A) Reverse transcription-polymerase chain reaction (RT-PCR) results for human Müllerian inhibiting substance type II receptor (MISRII) from human (a) myometrium, (b) myoma, (c) endometrium, and (d) adenomyosis. (B) Quantification of RT-PCR signals obtained using densitometric analysis of the signal product optical density×resulting band area.
GAPDH, glyceraldehyde 3-phosphate dehydrogenase.
2. Expression of MIS/AMHRII protein by immunohistochemistry
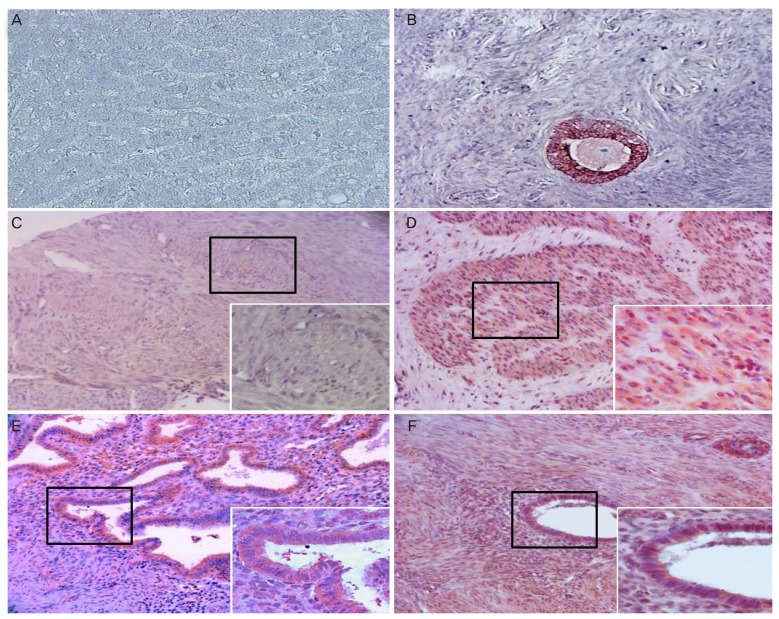
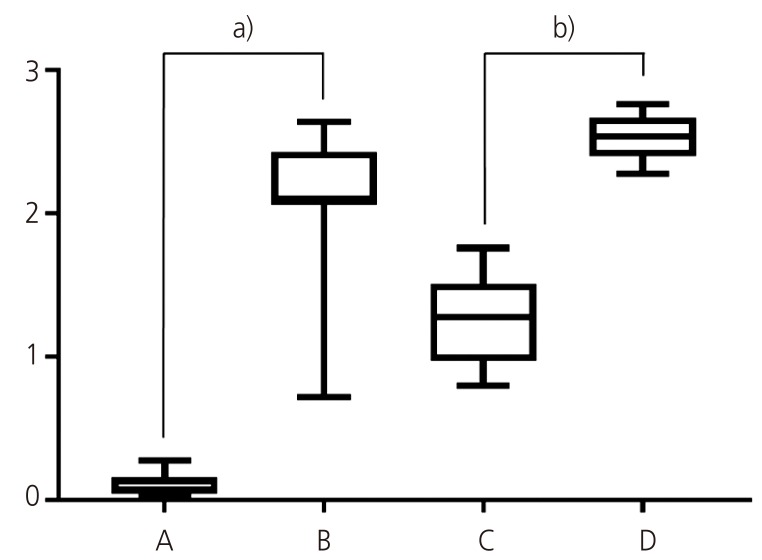
The frequencies of MIS/AMHRII expressing cells were 3.15%±1.69% in normal uterine myometrium, 51.95%±13.96% in uterine myoma, 31.10%±7.19% in endometrial tissue, and 64.65%±4.85% in adenomyosis tissue (Table 1). The most representative light micrographic findings of the myometrium, myoma, endometrium, and adenomyosis are presented in Fig. 2. The liver appeared to be MIS/AMHRII negative (negative control, Fig. 2A). The multiple layered granulosa cells of preantral follicle in ovary showed strong expression for MIS/AMHRII (positive control, Fig. 2B). Myometrium expressed little or no MIS/AMHRII (Fig. 2C), whereas in myoma, expression of MIS/AMHRII was strong (Fig. 2D). Endometrium showed moderate expression for MIS/AMHRII (Fig. 2E), but adenomyosis showed strong expression (Fig. 2F). The mean intensity scores for MIS/AMHRII expression were 0.13±0.07, 2.08±0.56, 1.24±0.29, and 2.59±0.19 in uterine myometrium tissue, uterine myoma endometrial tissue and adenomyosis tissue, respectively (Table 1, Fig. 3). MIS/AMHRII expression was higher in uterine myoma and adenomyosis tissues than that in normal uterine muscle and endometrial tissue, respectively.
Table 1
Frequency and intensity of Müllerian inhibiting substance/anti-Müllerian hormone type II receptor (MIS/AMHRII) in myometrium, myoma, endometrium, and adenomyosis

| Pathology | Myometrium | Myoma | Endometrium | Adenomyosis |
|---|---|---|---|---|
| Frequency (%) | 3.15±1.69 | 51.95±13.96 | 31.10±7.19 | 64.65±4.85 |
| Intensity score | 0.13±0.07 | 2.08±0.56 | 1.24±0.29 | 2.59±0.19 |
Fig. 2
Light photomicrographs of human (A) liver (negative control), (B) granulosa cells of preantral follicle in the ovary (positive control), (C) myometrium, (D) myoma, (E) endometrium, and (F) adenomyosis. Each image shows the expression of Müllerian inhibiting substance/anti-Müllerian hormone type II receptor (MIS/AMHRII) in the cell membrane. Right lower boxed area is of higher magnification (×400). Chromogen, amino ethyl carbazol. Magnification (×200).
Discussion
MIS/AMH has long been known to be produced in immature Sertoli cells in male embryos and binds to MIS/AMH receptors in primordial Müllerian ducts to cause regression of female reproductive structures [11]. Similar to other members of the TGF-β family, MIS/AMH binds to a serine-threonine kinase receptor complex of type I and II. Type II receptor, which is the primary receptor binds the ligand. Activin receptor-like kinase (ALK)-2, ALK-3 and ALK-6 serve as type I receptor for MIS/AMH. Both types are required for MIS/AMH signaling [311].
Several studies have demonstrated that MIS/AMH not only plays a significant role in embryonic sexual development, but also causes the growth inhibition in cells expressing MIS/AMH receptors. Since most gynecologic tumors originate from Müllerian duct derived tissues and MIS/AMH causes regression of the Müllerian duct, it is expected to inhibit the growth of the gynecologic tumors via MIS/AMH receptor-mediated mechanism [311]. Furthermore, several lines of evidence suggest that MIS/AMH inhibits proliferation in tissues and cell lines of MIS/AMH receptor-expressing gynecologic malignancies such as ovarian cancer [121314151617], cervical cancer [18192021], and endometrial cancer [2223]. In vitro studies using human epithelial ovarian cancer cell line (OVCAR 8) revealed that MIS/AMH inhibits growth of epithelial ovarian cancer cells [1417]. Park et al. [13] reported that MIS/AMH inhibits growth of ovarian cancer by deregulating the Wnt signal pathway via the β-catenin interacting protein. Similarly, MIS/AMHRII is also present in normal cervical and cervical cancer tissues, and MIS inhibits proliferation of cervical cells in vitro [1920]. MIS/AMH-mediated growth suppression is also observed in the C33A cervical cancer cell line [21]. MIS/AMHRII is highly expressed in a human endometrial cancer cell line (AN3CA), where MIS/AMH functions as a tumor suppressor by regulating signaling pathways that could contribute to endometrial carcinogenesis [23].
Myoma is the most common benign tumor of the female reproductive tract and is a monoclonal tumor arising from the smooth muscle tissue of the uterine myoma [2526]. Patients usually do not complain of any symptoms, but typical symptoms can be excessive menstrual bleeding, anemia, and chronic pelvic pain, and some women may be subfertile [28]. Treatments for uterine leiomyoma include medications, surgery, and radiologically guided intervention. Of these, surgical treatment includes myomectomy and hysterectomy. Myomectomy has the advantage of preserving the uterus, but the remaining, tiny or deeply embedded leiomyoma is likely to be growing and shown later, requiring another surgery. Therefore, the most effective treatment for myoma is hysterectomy [29]. Medical treatments include levonorgestrel-releasing intrauterine devices, gonadotropin-releasing hormone analogues, selective progesterone receptor modulators, oral contraceptives, progestins, and danazol [29]. However, these treatments do not eliminate the tumor and the symptoms recur when treatment is discontinued [2829].
Adenomyosis is a disease in which the endometrial glandular and stromal cells are invaded locally or diffusely in the myometrium, and the uterus is generally enlarged [27]. Most patients are asymptomatic, but typical symptoms are abnormal uterine bleeding, dysmenorrhea, and infertility [30]. Medical treatments include a levonorgestrel-releasing intrauterine system and gonadotropin-releasing hormone analogues. Surgical treatment includes hysterectomy and conservative surgery [30]. Without any consensus until now, hysterectomy and levonorgestrel-releasing intrauterine devices are still the main stream treatment for adenomyosis [30].
Although uterine leiomyoma and adenomyosis are common diseases in women of childbearing age, the most definite treatment for both diseases is a hysterectomy [2930]. Thus, various treatments for preserving the uterus, such as uterine artery occlusion by embolization are emerging; however, it is unclear how they affect fecundity and pregnancy [29].
Uterine leiomyoma and adenomyosis are tumors arising from the uterus which originates from the Müllerian duct. Therefore MIS/AMH is expected to inhibit the growth of these benign tumors via MIS/AMH receptor-mediated mechanism. However, unlike ovarian cancer, cervical cancer, and endometrial cancer, little is known about MIS/AMHRII expression or the effect of MIS/AMH.
In this study, we examined MIS/AMHRII expression in uterine leiomyoma and adenomyosis by using RT-PCR and immunochemistiry. The level of MIS/AMHRII mRNA expression in uterine myoma was higher than that in normal myometrium (4.51±0.26 vs. 0.08±0.09; P<0.05) and it was also stronger in adenomyosis than that in normal endometrial tissue (6.84±0.20 vs. 1.63±0.06; P<0.05). Myoma and adenomyosis showed stronger immunohistochemical expression for the MIS/AMHRII protein also.
We have demonstrated that myoma and adenomyosis express the MIS/AMHRII. The presence of the MIS/AMHRII in these cells identifies them as potential targets for MIS/AMH. MIS/AMHRII expression in myoma and adenomyosis was stronger than that in normal myometrium and endometrium. Our results suggest that MIS/AMH may act on myoma and adenomyosis to inhibit MIS/AMH-mediated cell growth and that MIS/AMH can be used as a uterus preserving treatment for patients with myoma and adenomyosis.
This study is significant as it is the first research to show that myoma and adenomyosis can be targets of MIS/AMH. However, it is unknown whether the receptor is functional or not. Thus, in vitro and in vivo studies are required to further investigate whether MIS/AMH can actually inhibit growth of myoma and adenomyosis.
 XML Download
XML Download