

 PDF
PDF ePub
ePub Citation
Citation Print
Print



Introduction
Ovarian cancer is the most lethal malignancy of the female reproductive system. Cytoreductive surgery followed by platinum based chemotherapy as standard therapy results in only 70% of partial or complete response, however, most will relapse within 2 years due to chemoresistance [1]. Recurrent disease is typically less responsive to current chemotherapy regimens. One of the causes could be the development of resistance mechanism to the drugs through genetic and acquired characteristics [2]. The development of enhanced target therapy and chemosensitization strategy to overcome chemoresistant cells is crucial.
Cancer stem-like cells were first identified in acute myeloid leukemia and many studies have reported specific tumor markers of cancer stem like cell which present more chemoresistant and more aggressive characteristics [3]. Recent researches have demonstrated that CD44 expression in ovarian cancer cells defines a tumorigenic cell population that expresses stemness. CD44 is a surface transmembrance glycoprotein involved in cell-cell, cell-matrix interactions which affect cellular growth, differentiation and motility. Increased expression of CD44 is associated with poor prognosis and resistance to chemotherapy [4]. CD117, known as c-kit, is a receptor tyrosine kinase. Increased CD117 expression level was observed in ovarian cancers with possession of cancer stem-like cell properties including self-renewal, differentiation, a high tumorigenic potential and chemoresistance [5]. To verify the characterization of stem-like cells, gene expressions of specific genes such as octamer-binding trascription factor 3/4 (OCT3/4), nanog homeobox (NANOG), and sex determining region Y-box 2 (SOX2) are demonstrated. They are named as stemness genes and they play a significant role in developmental processes including embryogenesis, neurogenesis, stem cell expansion and hematopoiesis. They are key regulators for the formation and/or maintenance of inner cell mass and for self-renewal of pluripotent stem cells [6].
Salinomycin is a monocarboxylic polyether antibiotics isolated from Streptomyces albus. It is originally used as an agricultural antimicrobial agent in poultry and for nutrition. It promotes the mitochondrial and cellular potassium efflux and inhibits mitochondrial oxidative phosphorylation thus affecting mitochondrial and cell membranes [7]. Gupta et al. [8] first reported salinomycin as most effective agent against breast cancer stem cells in 2009. By selective inhibition of breast cancer stem cells in mice it could reduce the size of tumors and increase necrosis and apoptosis. Currently, it is reported that salinomycin can target cancer stem-like cells in various human cancers including gastric cancer, lung, osteosarcoma, colorectal cancer, squamous cell carcinoma and prostate cancer [9]. However, there are not many reports about the apoptotic effect of salinomycin on human ovarian cancer cell line. Thus with understanding of specific characteristics of cancer stem-like cell we can expect synergistic effects of salinomycin with classical chemotherapy regimen.
The aim of present study was to investigate the expression of CD44 and CD117 in cancer stem-like cells from ovarian cancer and to evaluate the effect of salinomycin on growth inhibition of ovarian cancer stem-like cells.
Materials and methods
1. Reagents and cell line
OVCAR3 cells were purchased from ATCC (Manassas, VA, USA) and were grown as a monolayer in RPMI-1640 medium supplemented with 100 units/mL penicillin/streptomycin and 10% (v/v) fetal bovine serum at 37℃ in a 5% CO2 humidified atmosphere. Salinomycin and paclitaxel were purchased from Sigma-Aldrich (St Louis, MO, USA). Fetal bovine serum, penicillin/streptomycin solution, trypsin, RPMI-1640 medium were from HyClone (Logan, UT, USA).
2. Isolation of cancer stem-like cells
Tumor sphere culture was performed in ultra-low attachment dishes (Corning, Corning, NY, USA), supplemented with 5 µg/mL insulin (Sigma-Aldrich), 20 ng/mL human recombinant epidermal growth factor (Invitrogen, Carlsbad, CA, USA), 10 ng/mL basic fibroblast growth factor (Invitrogen) and 0.5% bovine serum albumin (Sigma-Aldrich) [10]. The CD44+CD117+ cells were sorted from the OVCAR3 cell line by using the magnetic-activated cell sorting (Miltenyi Biotec, Bergisch Gladbach, Germany). First, CD44+ subsets were isolated by using mouse antihuman CD44 antibody coupled to magnetic microbeads (Miltenyi Biotec), followed by the magnetic column selection or depletion. Second, resulting cells were then depleted of CD117- subsets by using mouse antihuman CD117 antibody coupled to magnetic microbeads (Miltenyi Biotec), and CD44+CD117+ cells were named as cancer stem-like cells (OVCAR3 CD44+CD117+).
3. Immunofluorescence
Cell suspensions were washed with phosphate buffered saline (PBS), smeared on clean slides and air dried prior to immunofluorescence microscopy. Slides were incubated with CD44 anti-mouse monoclonal antibody (Abcam, Cambridge, UK) diluted at 1:100 with PBS for 30 minutes at room temperature. Then the smear was incubated with Alexa Fluor 488 Goat anti-mouse IgG (Life Technologies, Carlsbad, CA, USA) diluted at 1:1000 with PBS for 30 minutes at room temperature. The preparations were washed again and the DNA was counter-stained with Hoechst 33342 (Sigma-Aldrich). Immunofluorescence microscopy was performed using an inverted microscope (DMIRB, Leica Microsystems, Wetzlar, Germany) equipped with ×100 fluotar objective and appropriate filter sets for UV and combined blue/green excitation.
4. Flow cytometry
Dissociated cells were counted and transferred to a 5-mL tube, washed twice with PBS, counted and resuspended in PBS at 1×106 cells/100 µL. Then, the antibodies allophysocyanin anti-human CD117 and fluorescein isothiocyanate anti-human CD44 (Becton Dickinson, San Jose, CA, USA) (each at a dilution of 1:40) were added and incubated for 20 minutes on ice in dark. The respective isotype control antibodies were used at the same concentrations according to the manufacturer's instructions. After being washed twice with PBS, samples were resuspended in 500 µL PBS and analyzed on a flow cytometer (FACS Calibur, Becton Dickinson).
5. Reverse transcription polymerase chain reaction
Total cellular RNA was extracted by using the TRIzol reagent (Invitrogen) according to the manufacturer's protocol. Reverse transcription polymerase chain reaction (RT-PCR) was used to analyze the expression of mRNA for OCT3/4, NANOG, SOX2, and glyceraldehyde 3-phosphate dehydrogenase (GAPDH) as internal control. The respective primers (Bionics, Seoul, Korea) and conditions were as follows: for OCT3/4, sense 5'-GAC AAC AAT GAA AAT CTT CAG GAG A-3' and antisense 5'-TTC TGG CGC CGG TTA CAG AAC CA-3' (annealing at 62℃, 40 cycles); for NANOG, sense 5'-ATG CCT CAC ACG GAG ACT G-3' and antisense 5'-CTG CGT CAC ACC ATT GCT A-3' (annealing at 60℃, 35 cycles); for SOX2, sense 5'-CAT GTC CCA GCA CTA CCA GA-3' and antisense 5'-GGG TTT TCT CCA TGC TGT TT-3' (annealing at 62℃, 40 cycles); for GAPDH, sense 5'-GCC AAG GTC ATC CAT GAC AAC-3' and antisense 5'-GTC CAC CAC CCT GTT GCT GTA-3' (annealing at 60℃, 25 cycles). Amplified products were separated by electrophoresis on 1.5% agarose gel and digitized by the Chemi-doc XRS imaging system (Bio-Rad, Hercules, CA, USA).
6. Protein isolation and immunoblotting
Cell (1×107/mL) extracts were prepared in 1× RIPA buffer (Thermo Scientific, Rockford, IL, USA) containing protease and phosphatase inhibitors with EDTA (Thermo Scientific, Rockford, IL, USA). Protein concentrations were measured using Bio-Rad Protein Assay Reagent (Bio-Rad) following the manufacture's protocol. Aliquots of protein were separated by 10% SDS-PAGE and transferred to nitrocellulose membrane (Millipore, Bedford, MA, USA). The membrane was blocked with Tris buffered saline containing 5% skim milk and 0.2% Tween 20 (Amresco Inc., Solon, OH, USA). After being washed, the membranes were incubated with appropriate primary antibodies (Cell Signaling, Beverly, MA, USA). The blots were reacted with horseradish peroxidase-conjugated secondary antibodies (Santa Cruz Biotechnology, Santa Cruz, CA, USA) and detected by the Super Signal (Thermo Scientific). The density of respective bands was analyzed by the Chemi-doc XRS imaging system (Bio-Rad). The data were presented as % of controls.
7. Cell viability assay
The number of viable cells exposed to paclitaxel and salinomycin was evaluated by a colorimetric 3-(4,5-dimethylthiazol-2,5-diphenyl tetrazolium bromide (MTT) assay. Initially, cells were seeded at a density of 5×104 cells/mL in 24-well plate, and then cultured for 24 hours to allow their adhesion to the plate. After pre-incubation, the culture medium was changed to experimental medium supplemented with paclitaxel (1, 10, 100, and 200 nM) and salinomycin (0.1, 0.5, 1, and 5 µM) alone or in combination for 48 hours. The intensity of the purple color formed by this assay is proportional to the number of viable cells. MTT reagent was added and incubated for an additional 4 hours at 37℃. The optical density was measured at 540 nm. The mean value and their standard deviation were calculated from triplicate experiments.
8. Statistical analysis
The data are presented as mean±standard deviation. Statistical analysis was conducted using one-way analysis of variance followed by Duncan's multiple range test for post hoc comparison by SPSS ver.17.0 (SPSS Inc., Chicago, IL, USA). Statistical significance was set at P<0.05.
Results
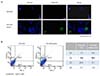
1. Expression of CD44 in ovarian cancer stem-like cells
The OVCAR3 cells were enriched for cancer stem cells by growing them as sphere forming conditions in ultra-low at-tachment dishes, the number of CD44+ cells increased (Fig. 1A).
2. Expression of CD44 and CD117 in OVCAR3 cells before and after sphere forming culture
To confirm the existence of the CD44+CD117+ phenotype in OVCAR3 cells, we cultured the cells by sphere forming method and demonstrated that the proportion of cells expressing CD44 and CD117 was 24.7% and 13.4%, respectively (Fig. 1B).
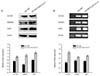
3. Expression of cancer stem cell genes and related proteins
The expressions of ovarian cancer stem cell genes and related proteins, including OCT3/4, NANOG and SOX2, was examined at the transcriptional and translational levels. Western blot data showed that the expressions of the OCT3/4, NANOG and SOX2 proteins were upregulated in OVCAR3 CD44+CD117+ cells compared with those in parental cells (Fig. 2A). The levels of OCT3/4, NANOG and SOX2 transcripts were also increased in sphere forming cells, and magnetic-activated cell sorting OVCAR3 CD44+CD117+ cells (Fig. 2B) compared with parental cells, as assessed using RT-PCR analysis.
4. Decrease in cell viability following salinomycin treatment
To evaluate the effect of salinomycin on ovarian cancer cells, OVCAR3 parental and OVCAR3 CD44+CD117+ cells were grown in the presence of different doses of paclitaxel and salinomycin. As shown in Fig. 3A, OVCAR3 CD44+CD117+ cells showed more resistance to paclitaxel than OVCAR3 cells. In contrast to control, salinomycin reduced the viability of OVCAR3 and OVCAR3 CD44+CD117+ cells in a dose-dependent manner (Fig. 3B).
5. Inhibitory effect of paclitaxel combined with salinomycin on ovarian cancer stem-like cell poliferation
The potential of salinomycin to inhibit the growth of ovarian cancer cell line (OVCAR3) and ovarian cancer stem-like cells (OVCAR3 CD44+CD117+) were determined by the MTT assay. Paclitaxel (10 nM) alone inhibited growth of OVCAR3 and OVCAR3 CD44+CD117+ cells by 19% and 5%, respectively. Inhibition of growth of OVCAR3 and OVCAR3 CD44+CD117+ cells by paclitaxel combined with salinomycin (0.1 µM) were 65% and 55%, respectively (Fig. 3C).
Discussion
Ovarian cancer is the fourth highest cause of female cancer death and the most common cause of gynecological cancer-associated death [11]. Although a significant proportion of women attain complete response with present standard management, approximately 70% will develop recurrence within 2 years.
Current evidence suggests that cancer stem-like cells are the putative mediators of chemotherapy resistance and tumor progression [1213]. A recent American Association for Cancer Research workshop defined cancer stem-like cell as a malignant cancer cell with a stem cell phenotype. Five criteria have been established to define the existence of cancer stem-like cells: self-renewal, minor population in tumor, reproducible tumor phenotype, multipotent differentiation into nontumorigenic cells, and expression of distinctive cell surface markers [141516]. The existence of cancer stem-like cells might explain why standard chemotherapy may kill tumors, however, spare cells that can give rise to a new tumor and cause relapse. Bapat et al. [17] reported the identification of cells with the characteristics of cancer stem-like cells in ovarian malignancy. Therefore, it is necessary to isolate and define tumoriogenic characteristics of cancer stem-like cells.
Recent researches have identified CD44 and CD117 as potential markers for the identification of cancer stem-like cells in ovarian malignancy [16]. Tumor spheroids generated from the ascite of ovarian malignancy patients are shown to have highly enriched CD44 and CD117 expressions. The CD44+CD117+ spheroid cells were resistant to chemotherapy and were able to initiate and serially propagate tumors in mice [18]. From analysis of 147 cases of epithelial ovarian cancers, Alvero et al. [19] found that the expression of CD44 was higher in metastatic tumors and tumor ascites. Subsequently, Luo et al. [20] reported that CD117+ lineage cells were capable of serial transplantation, asymmetric division, and the presence of these cells was correlated with chemoresistance. Significant association of CD117 expression with chemoresistance suggests that CD117+ ovarian tumor cells function as cancer stem-like cells. Zhang et al. [16] isolated ovarian tumor cells and cultured self-renewing spheroids. The spheroids were able to survive multiple passages, were resistant to platinum and paclitaxel and had the capacity to form tumors in immnuocompromised mice. Dual CD44+CD117+ cells were markedly more tumorigenic than their negative counterparts. In this study the OVCAR3 cells were enriched for cancer stem cells by growing them as sphere-forming conditions in ultra-low attachment dishes, and cultured spheroids were examined for expression of surface markers. The number of CD44+ cells increased. CD44 may become a possible therapeutic target for highly aggressive malignancies, including ovarian cancer.
Ovarian tumor spheroid cells overexpress stem cell genes. After demonstration of sphere-forming, RNA from sphere-forming cells and OVCAR3 CD44+CD117+ cells was analyzed for OCT3/4, NANOG, and SOX2. These genes are pluripotent transcription factor and have been identified as stem cell markers. Recently many studies reported that these factors are frequently over-expressed in aggressive poorly differentiated human tumors and play a significant role in developmental processes including embryogenesis, neurogenesis, stem cell expansion and hematopoiesis. They are key regulators for the formation and/or maintenance of inner cell mass and for self-renewal of pluripotent stem cells [6]. In our study sphere-forming cells and OVCAR3 CD44+CD117+ cells overexpressed stem cell surface marker and stemness gene compared with parental cells, which demonstrated that these cells contain stem-like cell characteristics.
Recently various compounds and drugs that selectively target cancer stem-like cells have been discovered. These agents include microbial-derived and plant-derived biomolecules, small molecule inhibitors targeting key components of intrinsic signaling pathways of cancer stem-like cells, antibodies directed against cancer stem-like specific cell surface molecules and some classical drugs that have been used for benign disease [21]. In particular salinomycin have been demonstrated to induce apoptosis and overcome apoptosis resistance in human cancer cell [22], suggesting possibility of salinomycin as an anticancer drug. Recent study by An et al. [23] demonstrated that salinomycin possesses anti-tumor activity and inhitibts breast cancer stem-like cells via an apoptosis-independent pathway and it downregulates NANOG, OCT4, and SOX2. In addition, a study in China showed that salinomycin can suppress the metastasis and invasion of bladder cancer cells by inhibition of epithelial-mesenchymal transition [24]. In present study, growth inhibitory effects of salinomycin were evaluated. Salinomycin reduced the viability of OVCAR3 and OVCAR3 CD44+CD117+ cells in dose dependent manners. Consequently, the combination of paclitaxel and salinomycin was applied in order to evaluate inhibitory effect on ovarian cancer stem-like cells proliferation. Compared to paclitaxel alone, combination of paclitaxel and salinomycin significantly reduced cancer stem-like cells proliferation. This study has a limitation because it only describes a phenomenon of salinomycin, not mechanism or pathway of salinomycin. To prove definite and reliable effect of salinomycin, further study including mouse model and in vivo study are necessary in the next step.
Although the standard combination of surgery and chemotherapy can effectively reduce tumor mass, most patients with residual ovarian cancer stem like cells eventually acquire chemoresistance. An ideal agent should be able to selectively target cancer stem like cells over normal cells. In the present study, the author identify expression of CD44+CD117+ surface markers in ovarian cancer which present specific tumorigenic characteristics. The potential of salinomycin to inhibit the growth of ovarian cancer was evaluated and salinomycin reduced the viability of OVCAR3 and OVCAR3 CD44+CD117+ cells in dose dependent manner.
This study demonstrates possibility of salinomycin as a novel and selective therapy for cancer stem like cells in ovarian cancer. These data contribute to development of diagnostic and therapeutic strategies in ovarian cancer.

 XML Download
XML Download