

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
The incidence of debilitating bone fractures increases with age, and as the aging population expands in nations around the world the public health burden of these age-related bone fractures continues to grow. The risk of hip fracture doubles every 5 to 6 years after age 60, underscoring the dramatic increase in fracture risk that accompanies older age.[1] Older individuals who are at greatest risk of fracture frequently show a spectrum of features broadly categorized as “frailty”, defined by unintentional weight loss, self-reported exhaustion, muscle weakness, slow walking speed, and low physical activity.[2] The functional and behavioral aspects of frailty such as muscle weakness and reduced walking speed are, not surprisingly, also associated with loss of lean mass in the form of sarcopenia.[3] The muscle weakness and reduced muscle mass that characterize sarcopenia are associated with an increased risk of falling, which is in turn a primary cause of bone fractures.[4] Thus, while middle-age is often associated with increased body weight in many developed countries, later life is frequently characterized by loss of body mass, loss muscle mass, bone loss, and ultimately bone fractures.
The constellation of factors that characterizes the frailty phenotype suggests that some of these features may be physiologically related through endocrine or paracrine pathways. The integrative physiology of exercise provides a very clear example of the extensive crosstalk that can exist among different organs and tissues.[5] For example, muscle is now recognized as a source of circulating myokines that can be impacted by resistance training and physical activity,[67] fat is a source of secreted adipokines that can be modulated by weight loss and may effect a number of different organs and tissues,[8] and bone is now acknowledged to produce a circulating osteokine osteocalcin that can alter insulin sensitivity and exercise adaptation.[9] It is also becoming better appreciated that some of these pathways are significantly altered with aging, modifying the network for tissue crosstalk that exists in younger individuals.[10]
The cytokine-like hormone leptin is a classic adipokine that is secreted by adipocytes, increases with weight gain, and decreases with weight loss.[8] Additional studies have, however, shown that leptin is also produced by skeletal muscle [11121314] as well as bone cells,[15] and leptin receptors are abundant in these musculoskeletal tissues.[1617] The leptin receptor is highly conserved in vertebrates, and leptin functions as a growth factor for both muscle and bone early in life.[18] In addition, altered leptin signaling may play a key role in the aging process, as leptin resistance is known to occur in the brain of aging rodents.[19] Leptin levels are positively associated with longevity in centenarians,[20] and the ratio of leptin to adiponectin is positively correlated with muscle strength in older adults.[21] In addition, higher leptin levels are associated with a reduced risk of dementia in elderly people.[2223] Leptin signaling and changes in leptin sensitivity with age are therefore likely to contribute to age-related degeneration of multiple organs and tissues. This review summarizes the evidence for leptin's role in the maintenance of muscle and bone mass with aging, and suggests future directions for research aimed at defining the basic mechanisms of organ crosstalk linking age-related changes in musculoskeletal tissues.
THE ANABOLIC EFFECTS OF LEPTIN ON SKELETAL MUSCLE
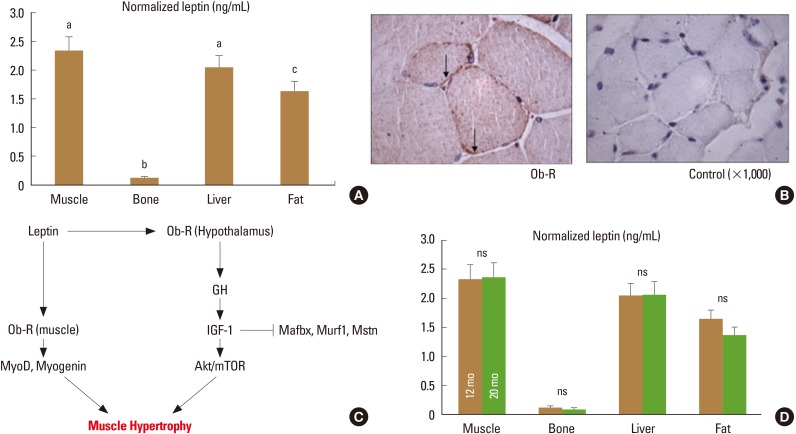
Leptin levels normalized for total protein are actually higher in mouse skeletal muscle than in mouse adipose tissue (Fig. 1A).[24] Muscle can accumulate fat, but fat does not accumulate substantially in normal mouse skeletal muscle, suggesting that the high concentrations of leptin in muscle are derived from muscle cells themselves.[25] This is further indicated by the observation that myoblasts secrete leptin in vitro,[12] and that leptin is released from skeletal muscle in vivo.[1314] Importantly, leptin release from skeletal muscle is only slightly less than release from adipose tissue (per unit tissue mass), yet muscle comprises a much greater percentage of body composition than fat, revealing that muscle is an important source of circulating leptin.[14] As noted above, leptin receptors are abundant in skeletal muscle (Fig. 1B),[17] their expression in skeletal muscle is altered with changes in physical activity,[1726] and the absence of functional leptin receptors in skeletal muscle impairs the capacity for myoblast proliferation and differentiation.[16] On the other hand, treatment of isolated primary myoblasts with leptin increases proliferation and the expression of myogenic genes (Fig. 1C).[16] These findings point to an autocrine function of muscle-derived leptin, but circulating leptin is also likely to have important anabolic effects on skeletal muscle. Leptin treatment increases muscle mass and decreases the expression of atrophy-related factors such as myostatin, muscle RING-finger protein-1 (MuRF1), and muscle atrophy F-box (MAFbx) in muscle (Fig. 1C).[1627] Circulating and muscle-derived insulin-like growth factor 1 (IGF-1) are both increased with leptin treatment in aged mice as well as in leptin-deficient mice, [2428] indicating that many of the anabolic effects of leptin on skeletal muscle are likely to be mediated by IGF-1. Finally, recent data suggest that leptin may also stimulate follistatin production in a circadian manner, and follistatin is a potent antagonist of myostatin.[29]
We have not detected a significant change in muscle-derived leptin with age (Fig. 1D), nor have we identified any changes in leptin receptor expression in skeletal muscle with age.[16] These observations are consistent with the finding that exogenous leptin can increase the expression of myogenic genes in primary myoblasts from aged mice,[16] and leptin treatment can increase muscle mass in aged mice and alter the expression of myogenic microRNAs such as miR-31 and miR-223.[30] In addition, leptin treatment produces a marked decrease in miR-489 expression in aged mice,[30] and miR-489 is known to maintain muscle satellite cells in a quiescent state.[31] Thus, leptin-induced suppression of miR-489 activity would be expected to enhance the capacity for muscle regeneration and repair in older animals. Although bone-derived mesenchymal stem cells appear to lose their responsiveness to leptin with age (see below), this does not seem to be the case in skeletal muscle. Age-related differences in the response of muscle and bone progenitor cells to leptin with age may also result from intrinsic differences in the progenitor cells themselves, as bone-derived stem cells appear to exhibit greater impairments with age compared to stem cells isolated from muscle or adipose tissue.[32]
LEPTIN AND AGE-RELATED CHANGES IN BONE MARROW CELL POPULATIONS
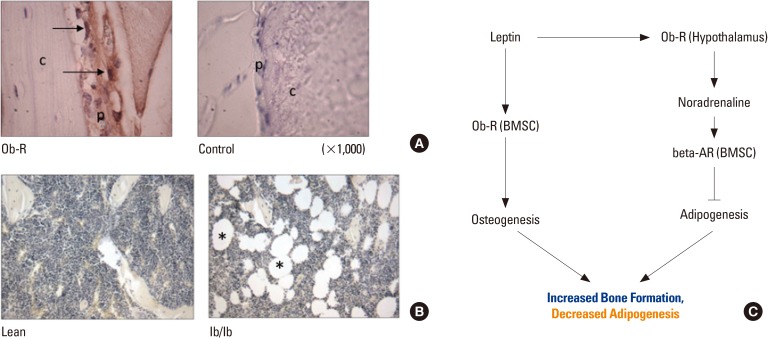
The central effects of leptin on bone, mediated by hypothalamic leptin receptors and the beta-adrenergic signaling network, were initially thought to suppress bone formation producing a low bone mass phenotype.[33] More recent studies show that the effects of leptin on the skeleton are quite complex, and that leptin deficiency is associated with low bone mass primarily due to reduced cortical bone.[343536] Central infusions of leptin in leptin-deficient ob/ob mice actually increase cortical bone formation and total bone mass,[28] but leptin also has important peripheral, direct effects on osteoblasts and bone-derived mesenchymal stem (stromal) cells (BMSCs). In fact, the leptin receptor is now regarded a marker of BMSCs[37] that mediates the switch between osteogenesis and adipogenesis.[3839] Leptin receptors are also abundant in the periosteum surrounding cortical bone (Fig. 2A). Leptin treatment of leptin-sensitive BMSCs increases the expression of bone morphogenetic protein 2 (BMP-2) as well as the secretion of the chemokine stromal cell-derived factor 1 (SDF-1) (CXCL12). [40] Other studies indicate that replacement of bone marrow cell populations with stem cells lacking the leptin receptor impairs bone formation, whereas restoring direct leptin signaling by transplanting cells expressing the leptin receptor enhances bone formation.[41] Moreover, as noted above, leptin can stimulate the growth hormone (GH)/IGF-1 axis, and IGF-1 is a potent osteogenic factor. Thus, leptin can effectively couple food intake and energy reserves with bone formation through both central and peripheral (direct) actions.
Aging appears to impact leptin's effects on bone metabolism in several ways. First, central leptin resistance, which is known to increase with aging,[19] is likely to attenuate leptin's central effects mediated by the hypothalamus. This may have a particularly important influence on bone marrow adipogenesis and the accumulation of bone marrow fat with age. Intrahypothalamic injection of leptin reduces marrow adipocytes in both mice[42] and rats[43], which appears to be mediated by beta-adrenergic signaling.[4244] Moreover, leptin deficiency due to either absence of leptin or calorie restriction increases marrow adipocytes (Fig. 2B).[45] These findings suggest that reduced hypothalamic sensitivity to leptin would be expected to support an overall increase in marrow adipocytes with aging (Fig. 2C). An increase in marrow fat with age-associated leptin resistance would also be consistent with the well-known phenomenon of increased marrow fat seen with advanced age in humans.
Second, evidence indicates that aging has a significant effect on BMSCs and their response to leptin. Individuals with osteoporosis have reduced levels of leptin in the bone marrow microenvironment,[46] BMSCs isolated from osteoporotic donors show lower leptin binding capacity than BMSCs from normal donors,[47] and leptin can suppress the adipogenic differentiation of BMSCs from healthy donors but not in BMSCs from osteoporotic donors.[48] The study by Astudillo and colleagues[47] suggests that leptin insensitivity due to decreased leptin binding occurs with age, but the molecular mechanisms underlying this phenomenon are not well understood. It is also unclear how leptin receptor expression, or the expression of microRNAs targeting the leptin receptor, are altered with age in BMSCs. BMSCs isolated from older individuals are observed to have impaired proliferation, increased senescence, and reduced potential for osteoblastic differentiation.[32495051] It is certainly possible that some of these age-associated changes in BMSC function documented in older donors may be associated with alterations in factors associated with leptin signaling, such as microRNAs targeting the leptin receptor or molecules such as suppressor of cytokine signaling 3 (SOCS3) and protein tyrosine phosphatase 1B (PTP1B) that inhibit leptin signal transduction.[5253]
DISCUSSION
Leptin secretion links food intake and energy reserves with energy expenditure, growth, and reproduction.[54] The role of altered leptin signaling in aging has, however, received less attention. Aging is associated with overall reduced calorie intake as well as reduced protein intake,[555657] which would be expected to lower circulating levels of leptin. Not surprisingly malnutrition in the elderly is in many cases associated with reduced leptin,[5859] as well as with reduced subcutaneous fat and reduced lean mass.[60] Many of the physical changes observed with older age such as reduced lean mass, bone loss, and cognitive decline may be related. For example, frailty and muscle weakness are associated with dementia,[6162] and aging and leptin resistance have been linked with the development of Alzheimer's disease.[63] These observations then raise the question of what interventions might increase leptin production and lean mass in older patients. We have found that the amino acid tryoptophan increases muscle-derived leptin, IGF-1, and follistatin in mice on a low protein diet,[64] and others have found that amino acid supplements plus exercise increase both serum leptin and musculoskeletal function in older adults.[65] These studies suggest that dietary interventions may provide one approach for reducing loss of lean mass with aging,[66] perhaps through a leptin-mediated pathway (Fig. 3). Recombinant human leptin (rhLep) has shown potential for increasing IGF-1 and markers of bone formation in women with hypothalamic amenorrhea,[67] although other studies indicate that the effects of rhLep on lean mass may be more modest.[68]
Resistance exercise can improve muscle strength and power in the elderly[6970] whereas bone is much less responsive to mechanical stimuli with age.[10] This may indicate that targeting muscle through nutritional supplementation (Fig. 3) as well as with resistance exercise is likely to have a greater impact on reducing the risk of falls and fractures than targeting bone itself; however, given that leptin can mediate the differentiation of bone marrow stromal cells directly and reduce bone marrow adipogenesis via its receptors in the hypothalamus, increasing leptin levels with increasing muscle mass may have some positive effects on the skeleton. These changes, in addition to the fact that leptin may prevent some neurodegenerative decline with aging, suggest that increasing muscle mass and strength in the elderly may have multiple, positive effects on the brain and the skeleton.

 XML Download
XML Download