

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Cardiovascular disease is a leading cause of morbidity and mortality in women. Cardiovascular diseases are less frequent in premenopausal women but the risk of cardiovascular diseases increases in the post-menopause estrogen deficiency. On the other hand, considering that the use of female hormones after menopause is likely reduce the risk of cardiovascular disease, it enables to assume the cardiovascular protective effects of estrogen.[1]
Cardiovascular protective effects of estrogen were presented as the results of direct action on the cardiac blood vessels with beneficial changes in lipid and lipoprotein.[2] And these mechanisms become the basic foundation of the "estrogen cardiovascular diseases hypothesis". Clinically important points standing out among the direct actions of estrogen on the blood vessels are the vasorelaxing action of estrogen by which estrogen expands blood vessels and increases the blood flow, while reduces the vascular resistance.
A so-called phytoestrogen, the plant originated estrogen is a material similar to the estrogen existing in the plants and essentially it is a natural phenolic compound, but as it presents structural or functional similarity to estradiol-17β, it can show in vivo estrogenic effects. Therefore, recently attempts have been made to use phytoestrogens in prevention and treatment of various symptoms and diseases that appear from estrogen deficiency after menopause as an alternative to the estrogen.[3]
Equol, as an in vivo metabolite of daidzein is natural non-steroidal polyphenols which is one class of the phytoestrogen, shows the best binding affinity with the estrogen receptor and the strongest oxidative actions among phytoestrogens. However, the equol has been known to be possible for its generation in about 30 to 50% of adult population, and the reason is presumed as due to the different intestinal microflora. And there has been reported that the one who can generate equol among those intake of phytoestrogen can obtain more clearer positive effect due to such appearance of equol.
Phytoestrogen including equol is assumed to present the similar actions in the cardiovascular system by having its structure and functions similar to those of estrogen, but in actuality, studies about it are lacking.
Vasodilation caused by estrogen is mainly controlled by a vasodilator substance, nitric oxide produced in the endothelium of blood vessels[4,5,6,7,8,9] and prostacyclin[10,11] or a vasoconstrictor substance, endothelin-I[12] and platelet-derived thromboxane.[13] However, as it was proved that the vasodilation activity was still existed in the blood vessels from which the endothelium was removed, assumption were also made that the vasodilation activity still existed by the endothelial-independent mechanism of the blood vessels.[13,14]
Calcium is one of the most important minerals in biological phenomena and in particular, it is an essential element for the regulation of contraction and relaxation of all muscles. Changes in intracellular calcium concentration of vascular smooth muscular cells regulate muscle contraction, which is induced by increase of intracellular calcium concentration in muscle cells. The concentration of the calcium ions in cytoplasm is controlled by endocytosis or exocytosis of calcium or by release or re-uptake of calcium into cytoplasm from intracellular pool of calcium. Influx of calcium ions into cells is mediated via calcium ion channels present on the cell membrane.
Calcium ion channels are categorized largely into voltage-dependent calcium channel[12,14,15,16,17] and receptor-dependent calcium channel[18,19] depending on the elements that determine intracellular access of calcium ions.
In addition, there is potassium channel existing in vascular smooth muscle cell membrane in addition to calcium channel, which causes hyperpolarization of the cell membrane so as to close calcium channel indirectly, resulting in the blood vessel relaxation.
If calcium ions are essential for the in the vascular smooth muscle contraction and the intracellular calcium ion concentration is to be regulated by calcium channel as well as potassium channel present in the cell membrane, it can be possible to presume the potential effects of equol as a phytoestrogen on the tonicity of vascular smooth muscle through these channels.
To this end, this study intended to investigate the effects of a phytoestrogen, equol on arterial tone of the endothelial cells-removed human uterine artery, using a uterine artery obtained during hysterectomy, and the study aimed to study the action of equol on the roles of calcium channel and potassium channel existing in the uterine artery smooth muscle cells when equol had effects on arterial tone.
METHODS
1. Study population
This study used the uterine artery obtained during hysterectomies performed in the Obstetrics and Gynecology Department of Chung Ang University Hospital as study material.
This study was conducted in 15 patients in total after exclusion of patients with past history of cardiovascular diseases or with risk factors for cardiovascular diseases affecting the blood vessels such as hypertension, diabetes and hyperlipidemia. The surgical indications of patients were 10 cases of uterine myoma, 2 cases of adenomyoma, 1 case of cervical dysplasia, 1 case of uterine prolapse, and 1 case of functional metrorrhagia. The age of the target group (mean±standard deviation) was 52.3±8.1 years and 11 of them were women before menopause, whereas 4 were women after menopause.
2. Preparation of uterine smooth muscle's sliced fragment
After carefully excised uterine artery from the uterus sample obtained via hysterectomy, immediately but gently rub the vascular endothelium using a wooden cotton swab, and removed it from the vessel. In the physiological solution saturated with oxygen for 100%, a sliced rectangular-shaped fragment of vascular smooth muscle (10 mm in length, 2 mm in width) was prepared while separating the connective tissues of the blood vessel wall by a micro scissors and the top rectangular-shaped smooth muscle (length 10 mm, width 2 mm) made fragment.
3. Measuring contraction and relaxation of uterine artery smooth muscle
After moving the vascular smooth muscle flap in the experimental container of 25 mL capacity, connet it to a isometric tension transducer (isometric force transducer 52-9545, Harvard, UK) and the contraction and relaxation of the vascular smooth muscle were measured by using a physiological recorder (physiograph, 50-8630, Harvard, UK). The preferred method of the test was the same as in previous tests.[20,21]
The test conditions were as given state of the resting tension of 2 g in a constant temperature of 37℃ and 100% oxygen was supplied. The surgery-induced damage was repaired while the contractile force was measured after resting for an hour or more, changing physiological saline as test solution in every 30 min in order to avoid the accumulation of metabolites.
The composition of the physiological solution was made with NaCl 140 mM, KCl 5 mM, CaCl2 2 mM, MgCl2 1 mM, and glucose 11 mM, and the pH was adjusted to 7.4 by using NaOH. All sampled chemicals including equol and those used in this test was purchased from Sigma Chemical Company.
Every test was repeated eight times and more, and the most representative illustration was used for the results.
4. Measuring the effect of equol on the contraction of the uterine artery smooth muscle
1) The effect of equol on the calcium channels of vascular smooth muscle cell membrane
(1) The effects of equol on the phenylephrine-induced contraction of the uterine artery smooth muscle
As α-adrenergic receptor stimulant, phenylephrine activates the receptor-dependent calcium ion channels. Phenylephrine 10-5 M was used to induce contraction of uterine artery smooth muscle flap as the default. By administration of equol for phenylephrine-induced contraction, its relaxation effects were observed. The equol was used with 6 different concentrations from 10-11 to 10-6 M, and equol was administered at 5 min intervals by each concentration and its relaxation effects were observed.
(2) The effects of equol for vasospasm caused by the high concentration potassium solution
A high concentration potassium solution directly activates the membrane potential calcium ion channels of vascular smooth muscle cell. In this study, a 70 mM potassium solution was used to induce the contraction of uterine smooth muscle flap, which was used as the default.
After administration of six different concentration equol to the blood vessel fragment which was induced by a high concentration potassium concentration solution of 70 mM at 5 min intervals in the same manner, the size change of the contractile force was recorded.
(3) Evaluation of contraction induced by phenylephrine and a high concentration potassium solution after pretreatment of equol
The study intended to evaluate the effects of equol, after pretreating uterine arterial smooth muscle with 10-8 M of equol first, and by inducing vasospasm with the six concentrations of phenylephrine from 10-9 to 10-4 M. In addition, it intended to evaluate the effects of pretreated equol after performing the same pretreatment and by inducing vasospasm with the four concentrations of high concentration potassium solution from 35 to 140 mM.
2) Effects of equol on potassium channel of vascular smooth muscle cell membrane
Two kinds of potassium channel antagonist, tetraethylammonium (TEA) and aminopyridine were used in studying the effects of equol on vasospastic response to the intervention of potassium channel of uterine arterial smooth muscle.
A the non-specific potassium (K+) channel blocker, TEA 10 mM was used for pretreatment at 30 min prior to administration of phenylephrine, and after induced vasospasm by phenylephrine 10-5 M, the six concentrations of equol were dosed and the relaxation effects were observed, then compared the effects to that in the control group. After inducing vasospasm with A-type potassium channel blocker, 4-aminopyridine 10-5 M at 30 min prior to induction of vasospasm with phenylephrine in the same manner, the six concentrations of equol were dosed and the relaxation effects were observed, then compared the effects to that in the control group.
As for the relaxation degree of uterine arterial smooth muscle, it was indicated as a percentage of the value obtained by dividing the tonicity at maximum relaxation in each concentration after administration of equol as well as the tonicity at maximum relaxation after administration of phenylephrine.
RESULTS
1. Measurement of the equol's effects on the uterine arterial smooth muscle
1) The effects of equol on calcium channel of vascular smooth muscle cell membrane
(1) The effects of equol on the vasospasm of the uterine arterial smooth muscle induced by phenylephrine
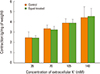
After removing the endothelium of uterine artery, and after the vasospasm was induced to the remaining uterine arterial smooth muscle fragments by phenylephrine 10-5 M, when administered equol with the concentrations from 10-11 to 10-6 M, the smooth muscle had shown relaxation response which was concentration-dependent according to the increase of concentration (Fig. 1).
(2) The effects of equol on inducement of vasospasm by a high concentration potassium solution
After the contraction of the uterine arterial smooth muscle by a high concentration potassium chloride solution 70 mM in the same way, when administered equol in the same concentration range (from 10-11 to 10-6 M), the contractile response of smooth muscle were not shown with any significant relaxation response (Fig. 2).
(3) Evaluation of vasospasm with phenylephrine and a high concentration of potassium solution after pretreatment of equol
After pretreatment of uterine arterial smooth muscle using equol 10-8 M first, and after induction of vasospasm using the six concentrations of phenylephrine from 10-9 to 10-4 M and the four of highly concentrated potassium solutions from 35 to 140 mM, the effects of pre-treated equol was evaluated.
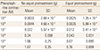
The vasospasm induced by the six different concentrations of phenylephrine from 10-7 M had shown concentration-dependent significant reduction in the group of pretreatment with equol (Fig. 3, Table 1), whereas vasocontraction by the other six concentrations of phenylephrine from 10-7 M showed a concentration-dependent significant reductions in the pretreated equol groups (Fig. 3, Table 1) but the vasospasm induced by the four of highly concentrated potassium solution did not have significant differences regardless of whether the pretreatment of equol at all concentrations of potassium solution (Fig. 4, Table 2).
2) Effects of equol on the potassium channel of vascular smooth muscle cell membrane
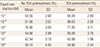
After a pretreatment of the uterine artery with TEA, the relaxation response of uterine arterial contraction caused by phenylephrine had shown no significant differences to the relaxation response from a single dose of phenylephrine in the control group according to the equol concentrations (Fig. 5, Table 3).
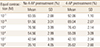
Also the relaxation response of equol for the uterine arterial contraction caused by phenylephrine after pretreatment with 4-aminopyridine did not show any significant differences to the response in the control group administered with equol that was not pretreated with 4-aminopyridine (Fig. 6, Table 4).
DISCUSSION
Cardiovascular disease is one of the main causes of mortality and morbidity in the female population of Korea as well as western countries. According to the observational studies,[22] risks of cardiovascular disease from hormone therapy, showed a decrease by about half, and a randomized, controlled study also reported that the risk was reduced in women ≤60 years of age or in early menopause stage of 10 years. The reduction in the risk of cardiovascular disease from hormone therapy is caused primarily by the improvement of lipids and lipoprotein, and the contribution of plasma lipids and lipoprotein to such reduction is about 30%. Although the non-lipid mechanism of estrogens is not clearly identified yet, it is suggested the direct action on the cardiovascular dynamics is more important. While the postmenopausal deficiency of female hormone leads to an increase in the vascular resistance, the blood vessels are dilated, vascular resistance is reduced, and the blood flow is increased from hormone replacement therapy.
Phytoestrogens as a natural phenolic compound derived from plants, are structurally and functionally similar to the endogenous estrogen. Therefore, phytoestrogens are expected to exhibit the actions of estrogen in the body and their effects on the cardiovascular system are assumed to be able to replace estrogen in this perspective.
This study is to investigate response of equol as one of phytoestrogens to cardiovascular vascular system, in particular, to vascular smooth muscle and its mechanism, and to observe whether it exhibits appearance similar to the endogenous estrogen. As far as the authors know, this study is considered as the first study to observe the effects of equol that is phytoestrogens by using human blood vessels.
From the results of this study, a phytoestrogen, equal causes endothelial-independent vascular relaxation response, and this response is mediated only through antagonism of calcium channels, without involving actions on potassium channels. As a phytoestrogen, equol exhibited concentration-dependent relaxation response to the contractile reactions induced by 10-5 M phenylephrine in the endothelial-removed human uterine artery, it had proved that equol might act as calcium antagonist. However, as equol did not demonstrate any significant relaxation response to the contraction induced by a high concentration KCl solution of 70 mM, the calcium antagonizing action of equol was not a phenomenon, which was induced by the mediation of the voltage-dependent calcium channels. So that it was assumed as action mediated by a receptor-dependent calcium channels. Relaxation response of equol to the phenylephrine-induced uterine arterial contraction did not show any significant variations by the pretreatment with high concentration tetramethylammonium and 4-aminopyridine, therefore, it was assumed that potassium channels was not involved in the vascular relaxation of equol.
Phytoestrogens are natural compounds originated from plants and show structural or functional similarity to estrogen and physiologically it is a substance having estrogen and anti-estrogen properties. Phytoestrogens can be chemically classified into 4 kinds of estrogen that are steroids, saponins, terpenoids and phenols. Double phenolic phytoestrogens is most abundant in the human diet, as these are the main types of phytoestrogens classified into seven subfamilies. Among the subfamilies, isoflavones, lignans and coumestans are the most important three groups. In isoflavones, biochanin, formononetin, genistein and daidzein have physiologically significant estrogenic activities, and the most critical isoflavones found in humans are genistein and daidzein. Phytoestrogens have been used for the relief of menopausal symptoms and prevention of osteoporosis and hormone-dependent cancers in women at menopausal period by replacing existing hormonal therapy, osteoporosis, and hormone-dependent cancer.
Equol is a metabolite of daidzein that is 7-hydroxy-3-chroman, heterocyclic phenol which was separated from the urine of pregnant horses for the first time by Marrian and Haslewood in 1932, and named equol as its chemical structure was identified. Equal has an estrogenic activity and as it was widely known as identified that it bound to receptors in the uterus and identified as the cause of infertility syndrome referred to as "clover disease". Since equol is not a natural constituent of plant, strictly speaking, it is not a phytoestrogen. Rather, equol can be defined as non-steroidal estrogen belongs to isoflavones as a metabolite entirely resulted by the metabolism of intra-intestinal bacteria.
The main dietary sources of equol are soy protein and soy bean foods in human because they are the most abundant sources of daidzin and daidzein that are precursors of equol. Equol is formed from daidzein but not from genistein.
Equol is the only metabolite that has the activity of the hormone among isoflavone metabolite, and it is very important because its estrogenic activity is as high as 5-fold of daidzein[23] and twice of genistein.[24] Genistein has a higher affinity for estrogen receptor-β whereas daidzein has a relatively lower affinity than genistein. The binding capacity of equol to human estrogen receptor α and β is 10 to 100 times greater than the precursor daidzein, and similar to genistein but the transfer by equol is more powerful than any other isoflavones. When observed the effects on uterine weight of the immature rat, equol had shown its estrogenic effects twice greater than genistein.[25]
Equol has the strongest antioxidant activity among all isoflavones. Although isoflavones are considered as weak antioxidants in vitro tests, but it has sufficient in vivo effects to inhibit lipid peroxidation. Inhibition of lipid peroxidation due to such superior antioxidant activity of equol, the equol producer is expected to reduce the risk of cardiovascular disease even more.
So far, the studies of soy beans have been conducted mainly focusing on genistein and daidzein. However, as the metabolic and physiological characteristics of equol as the metabolite of daidzein had been revealed, the importance of equol has been gradually increasing.
For cardiovascular system, whether phytoestrogen similar to estrogen can act as same as estrogen is not clearly identified. Furthermore the research of equol's effects on the cardiovascular system is so far very rare.
Cardiovascular protective effects of phytoestrogens found other than equol identified up to now are anti-atherosclerotic functions, which are assumed to be performed by the lipid and non-lipid mechanisms, likewise cardiovascular protective mechanisms of estrogen.
According to the studies that had observed changes of the blood lipid/lipoprotein level after ingestion of soybean or soy protein,[26,27] the target group that had shown improvement in lipid and lipoprotein metabolism was reported to be limited only to equol producer.
According to a 2-year study with subset analysis for the lipid in postmenopausal women, soybeans showed maximal cholesterol-lowering effects in the equol producer. Meaning, the total blood cholesterol level had shown a significant decrease of 7.2% from equol producer, whereas non-equol producer had shown insignificant decrease of 3.0% only.[26] Similar effects were observed for the spine bone mass. A study in middle age men and women with hypercholesterolemia or high blood pressure[27] also had presented that total cholesterol, low-density lipoproteins (LDL)-cholesterol, LDL:high-density lipoproteins (HDL)-cholesterol, non-triglyceride Lp (a) were significantly decreased only in equol producer, thus the improvement of blood lipids were presented as confined to equol producer.
As for non-lipid mechanism of equol that reduces the risk of cardiovascular disease, vasorelaxation is suggested likewise estrogen. According to several studies,[28,29,30,31,32,33] equol relaxes the blood vessels and it allows assumption that such action of equol is helpful for reducing the risk of cardiovascular diseases.
In mice, equol increases the discharge of urinary salt, and allows the aortic ring contracted by phenylephrine to be relaxed presenting vasodilation action.[30] A study conducted by Chin-Dusting et al.[29] also presented that equol had exhibited significant antagonistic action on the contraction of thoracic aortic ring in mice, which was induced by noradrenaline together with the metabolites of 17β-estradiol and the four isoflavones. When increased the concentration of equol to 0.1 µg/mL, 1 µg/mL, and 10 µg/mL, it did not inhibit the contractile responses by noradrenaline in 0.1 µg/mL but as it had shown effects of inhibition of 45.8% and 58.7% in 1 µg/mL and 10 µg/mL, respectively. And the inhibitory action was observed as dose-dependent. On the other hand, as it had shown 90.4% of inhibitory actions in 17β-estradiol 1 µg/mL, the effect of equol was assumed to be lower compared to estradiol at the same concentration.
In addition, in a study[31] that had observed vasodilatation of equol in the carotid artery and basal artery of brain in mice with normal blood pressure and those with hypertension, equol had shown vasodilation in the mice with normal blood pressure whereas it also maintained vasodilation induced by equol in the mice with angiotensin II induced hypertension. However, in case of daidzein, the precursor of equol, vasodilation was observed in mice with normal blood pressure, but vasodilation was significantly inhibited presenting patterns different from equal in the mice with hypertension.
In these three animal studies, equol commonly had caused vasodilation. From a human study, when allowed soy bean isoflavones containing diet in adults with hypercholesterolemia for 4 weeks, the equol producer had shown significant increase of brachial artery flow-mediated vasodilatation compared to the non-equol producer.[34] Although there were not many studies conducted so far on the mechanisms of equol for vasorelaxation, in the light of studies up to now, action of equol for vasorelaxation was identified as being resulted mainly from endothelial cell-nondependent vasorelaxation. Isoflavones increased the gene expression of endothelial cell nitric oxide synthase (eNOS), resulting in improvement of endothelial cell-dependent vasorelaxation and in lowering the blood pressure.[33]
Phytoestrogen, likewise estrogen, has classic mechanism mediated by transcriptional activation of estrogen-responsive genes in which the estrogen receptors are involved. Hormone-receptor complex can adjust an eNOS by causing transcription activity binding to specific estrogen response element (ERE) existing in the promoter region of the target gene. However, the activity of the target gene induced by estrogen can be mediated also by other transcription factors, for example, the independent Ap-1 and nuclear factor-kappaB (NF-κB) unrelated to ERE. From an experiment used swine pulmonary artery and human pulmonary artery endothelial cells,[28] Human immunodeficiency virus (HIV) protease inhibitor ritonavir (RTV) had significantly reduced the endothelium-dependent vasorelaxation by a endothelium-dependent vasodilator, bradykinin D in the pulmonary arterial ring and equol had reverses such concentration-dependent effects of RTV efficiently.
RTV has reduced vasoconstriction caused by U46619, a thromboxane A analogue, but such reduction was blocked by equol. When treated RTV in the pulmonary artery and pulmonary arterial endothelial cells, expression of eNOS was significantly reduced, whereas equol had effectively inhibited RTV-induced eNOS down-regulation. To this end, it is assumed that equol effectively protect vascular function by reducing the down-regulation of vasomotor dysfunction and eNOS.
According to recent studies, it was assumed that the cell surface receptor was involved in rapid response to phytoestrogens in endothelium and smooth muscle, likewise estrogen. The activity of cellular membrane-estrogen receptor stimulates signaling cascades, resulting in an increase of eNOS activity through the regulation of the transcription factor mediated by a kinase. Equol (1-100 nM) had caused rapid endothelial-dependent and nondependent relaxation in the aortic ring of mice and it activated eNOS rapidly within 2 min in human aorta and umbilical cord venous endothelial cells.
In a study used the human umbilical vein endothelial cells, when observed signaling pathway where equol was involved in the rapid activation of eNOS and generation of nitric oxide (NO), equol had phosphorylation of extracellular signal-regulated kinase (ERK)1/2 and PI3-K/Akt rapidly, resulting in phosphorylated eNOS that binds to HSP 90. Due to such binding, generation of NO was increased, resulting in vasorelaxation.[32] However, in a study of vasorelaxation effects of equol conducted by Jackman et al.,[31] the vasorelaxation effects of equol was observed as the same in males and females, and it showed identical relaxation effects before and after the removal of the endothelium was the same.
In the light of such results, it was assumed that vasodilatation action of equol was exhibited regardless gender and the presence/absence of the endothelium. Also it showed vasodilatation induced by equol had shown its effects regardless of the presence of the NOS inhibitor (L-NAME). That is, even if the relaxation response of equol inhibits NOS using L-NAME, from the fact that it did not affect the relaxation response of equol, thus it was to determine the findings that the effects of equol do not depend on the generation of NO by endothelial NOS and such findings were consistent with the finding that it was endothelial-independent action.
In this study, we observed vasoconstriction and vasorelaxation using a denuded blood vessel, and it was considered as direct effects on the vascular smooth muscle. Calcium ion is an essential component for muscular contraction and if the intracellular concentration of calcium ion is elevated, it causes contraction whereas if the concentration of calcium ion is lowered to less than 10-8 M, it will cause relaxation. The contraction of vascular smooth muscles is caused by an increase in the phosphorylation of myosin and/or by an increase in protein-myosin interaction induced by phosphorylation of proteins associated with actin, that is, caldesmon or calponin. Increase in phosphorylation of myosin is realized by activity of myosin light chain kinase or down-regulation of myosin light chain phosphatase. Among them, the more important action, that is, activity of myosin light chain kinase activity is dependent on intracellular calcium ions. This means, an increase in intracellular calcium ion forms the calcium-calmodulin complex and activates protein kinase that is a protein phosphorylation enzyme, which leads to activation of myosin light chain kinase of the smooth muscle, ultimately resulting in phosphorylation of myosin. The phosphorylated myosin activates interaction with actin and causes contraction of the smooth muscle. The calcium concentration in the cytoplasm increases by inflow from outside of the cell, or by release from sarcoplasmic reticulum, an intracellular reservoir of calcium. In smooth muscle, calcium ions inflow into the cell by various stimuli such as active voltage, hormones, and vasoconstrictors, and calcium channels existing in the cell membrane are largely classified into two types, in accordance with the factor determining the entry of calcium ions. The calcium channels are activated by depolarization of the cell membrane so called voltage-dependent calcium channels,[12,14,15,16,17] and the channels being activated as hormone or vasoconstrictor combines with receptors are so called as receptor gated calcium channels.[18,19] In addition, there is such as an extension gated calcium channel which shows increase of the intracellular calcium influx is increased by extension of the smooth muscle.
In this study, phenylephrine was used for contraction of the vascular smooth muscle, which activates the α1-adrenergic receptor so as to open the receptor gated calcium ion channel and causes contraction of the vascular smooth muscle. In this study 10-5 M of phenylephrine was used to cause contraction to the estimated degree of about 70% of the maximum vasospasm caused by phenylephrine, and equol had shown a concentration-dependent relaxation for the vasospasm induced by 10-5 M phenylephrine. In addition, equol was pretreated with 10-8 M thereafter an induced contraction according to the concentration of phenylephrine was observed. The group with pretreatment with equol had shown significant reduction of contraction at the concentration at 10-7 M or higher. From these results, it was assumed that equol had shown calcium antagonism through the vascular smooth muscle receptor gated calcium channel, and such action of equol to antagonize calcium was assumed as concentration-dependent.
High concentration potassium solution is also used in this study to keep the concentration of extracellular potassium ions at higher concentration than the normal physiological state, and by introducing high concentration potassium ions into the muscle cells to stimulate depolarization of the cell membrane so that the depolarization may activate voltage-gated calcium channel, resulting in vascular smooth muscle contraction. That is, a high concentration potassium solution is a representative substance that activates the voltage gated calcium channels of the cell membrane. This study had shown that the smooth muscle contraction was not influenced by a high concentration potassium solution, when dosed the six concentrations of equol after induced the contraction of uterine arterial smooth muscle with a high concentration KCI 70 mM. In addition, there was no change observed at all KCl concentrations, according to pretreatment performed or non-performed, when the contraction was induced by 35 to 140 mM of KCl after pretreatment with equol performed. These results were assumed as evidences that had shown equol did not mediate calcium antagonistic actions through voltage gated calcium channels.
The results of this study, which had investigated the mechanism of action of equol through the calcium channels in vascular smooth muscle, were similar to the results of previous studies on a different phytoestrogen genistein.[21] However, when induced vasospasm by highly concentrated KCI using a bovine pulmonary artery, genistein had reduced contraction by about 50%, whereas daidzein was lesser effective than genistein, and equol did not show effects,[35] and from such results, the researchers addressed the importance of role of tyrosine phosphorylation that was inhibition of tyrosine kinase activity.
In the smooth muscle cell membrane, such as potassium channels exist in addition to calcium channels. This potassium channel mediates contraction of smooth muscle indirectly. The potassium channel present in the cell membranes of vascular smooth muscle allows potassium ions selectively to pass through the membrane and it acts as antagonist for activity of sodium channel and calcium channels in the regulation of excitability of the cells. The activity of potassium channel increases the release of potassium ions, resulted in hyperpolarization of cell membranes. Then hyperpolarization closes the voltage-dependent calcium channels so as to relax the blood vessels. Conversely, if potassium channel is closed, calcium channel is opened causing vasoconstriction.[36]
It has been identified as the four different potassium channels exist in the cell membranes of vascular smooth muscle, and estrogen and genistein cause vasodilatation by activating potassium channels.[37]
In this study, in order to observe the effects of equol on potassium channels, a nonspecific potassium channel inhibitor, 10 mM tetramethylammonium or 4-amino-pyridine an A-type potassium channel inhibitor were pretreated, the effects of equol of various concentrations on the induction of vasoconstriction by phenylephrine were observed. As the results, they had not caused any significant variation and consequently it enabled to assume that potassium channel did not involve in the vascular relaxation effects of equol. This result is different from the results of previous studies on a representative phytoestrogen genistein[21] and these findings enabled to assume that there were differences in effects on potassium channels depending on the type of phytoestrogens. Conclusively, the phytoestrogen equol acts antagonistically on calcium channels in the smooth muscle cell membrane, in particular, on the receptor-dependent calcium channels. But it is assumed that such did not affect voltage-dependent calcium channel or potassium channel and such calcium antagonistic action may cause endothelial-dependent vasodilatation. Because of such mechanism of action, it is expected that equol contributes potentially in the cardiovascular protective effects.
An isoflavone intermediate metabolite of isoflavone, dehydroequol also showed a vasodilatation action.[29,38] Chin-Dusting et al.[29] had observed the ability to inhibit the contractile responses by noradrenaline of dehydroequol for the first time. An isoflavone metabolite dehydroequol exhibited the strongest effects among all isoflavone metabolites. Dehydroequol had shown stronger action than equol, presenting actions similar to the action of 17β-estradiol. Such vasodilatation effects were not inhibited by a NOS inhibitor N-nitro-L-arginine (NOLA) or by a guanylate cyclase inhibitor 1H-1, 2, 4 oxadiazolo 4, 3-aquinoxalin-1-one, or by indomethacine. These findings were assumed as evidences showing that NO or prostacyclin may not play the main role in vasodilatation action of dehydroequol. However, in the studies conducted in use of human upper extremity arteries, dehydroequol had exhibited significant inhibitory effects for inhibition of NOS together with dose-dependent increase in blood flow. Judging from these results, it showed that isoflavones and metabolites including equal may represent different effects depending on the species and the type of blood vessels. According to recent studies, not only equol itself is important but also in fact the state that can produce equol is suggested as improvement factor of the cardiovascular system.[39] After a 7 day intake of 52 g of soy protein containing isoflavones 112 mg daily, when defined the one whose equol concentration is increased more than four times as the equol producer, depending on the blood equol generation ability, the equol producer in postmenopausal women who use tibolone had shown significantly lower arterial stiffness compared to non-producer and significant improvement of endothelial function. However, when dosed soybean additionally, there had been no effects on arterial stiffness and endothelial function. Given such phenomena, the ability to produce equol can be an independent vascular health marker presumably, and these findings are consistent with the results of previous study suggested the equol producers had shown relevance with lower blood pressure.[40]
Equol may be broadly classified as polyphenol, and by having hydrogen donor property due to a large number of electrons, it can clean free radical. In general, isoflavone had increased the expression of antioxidant genes.[33] In vitro antioxidant activity of equol has been well verified, and had shown the strongest antioxidant activity of all isoflavones. In the brain basal artery of a mouse, equol had exhibited its antioxidant action by reducing the production of superoxide induced by NADPH significantly,[31] and a HIV protease inhibitor, RTV had increased the production of superoxide anion significantly, but equal had reversed such effects of RTV,[28] and it was addressed that vascular function could be protected by such reduction of oxidative stress.
So far there had been no in vivo study about to which degree equol had effects on human lipid oxidation. Nevertheless, it was proved that antioxidant effect was suppressed by the inhibition by anti-oxidative effects from mediation of the oxidation of LDL cholesterol by cultured monocytes/macrophages through inhibition of superoxide radical generation by equol.[41] Due to such anti-oxidative action of equol, the equol producer is expected to reduce the risk of cardiovascular disease. Further researches will be needed to identify the vascular protection from calcium antagonistic and antioxidant effects of equol as non-lipid mechanisms.






 XML Download
XML Download