

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Salivary glands are composed of acinar cells, either serous or mucous, and ductal cells of several varieties. The primary saliva is formed in the lumen of the acinar cells, where the accumulation of ions generates a transepithelial osmotic gradient driving water movement. In the second stage of secretion, sodium is actively reabsorbed while the primary saliva passes through the duct. Therefore, alterations of sodium transporters and water channels may affect the formation and the ductal modification of the saliva.
Immunofluorescence labelling and confocal microscopy revealed a polarized distribution of Na+,K+-ATPase, the secretory isoform of Na+/K+/2Cl- cotransporter (NKCC2), and type 1 Na+/H+ exchanger (NHE1) in the basolateral membranes of acinar and intralobular duct cells1). Na+,K+-ATPase produces an inward-directed sodium chemical gradient in the acinar cell2). Although sodium influx is negligible in resting cells, it is rapidly increased by activation of NKCC2, NHE1, and nonselective cation channels in the acinar cell3). Blockade of NKCC2 causes significant reduction of acetylcholine-stimulated secretion4), in which the residual secretion is attributed to the operation of Na+/H+ and Cl-/HCO3- antiport5, 6). The amiloride-sensitive epithelial sodium channels (ENaC) are localized to the surface epithelial cells of the secretory duct. Among their three homologous subunits, only α-subunit can produce an amiloride-sensitive current7), while β- and γ-subunits are not functional on their own8).
On the other hand, there have been known multiple isoforms of aquaporin (AQP) water channels in the salivary gland. Among them, AQP5 may be the only isoform of AQP playing a major role in the salivary secretion process. Strong AQP5 labelling is located in the apical membrane of serous-type acinar cells from the rat submandibular gland9). The transepithelial water movement occurs through the apical AQP5 channels and paracellular pathways10), leading to the secretion of isotonic primary saliva. Although the presence of AQP1 and AQP8 has been also generally accepted, their role in the salivary secretion remains controversial11).
Over the last two decades, nitric oxide (NO) has been found to play a role in the regulation of various physiological functions. It has been also implicated in mediating the nerve-evoked vasodilator and secretory responses in the salivary gland. Parasympathetic nerve activity increases the submandibular blood flow, and elicits the flow of saliva and output of protein by mechanisms that involve in situ generation of NO in the ferret12). A previous study revealed that NO synthase (NOS) is not present in the acinar cells, but in neural terminals within the gland and in the apical membrane of the excretory and striated ducts, the cytoplasm of granular convoluted tubules, and, to a lesser extent, in the cytoplasm of excretory and striated ducts13). However, later investigators demonstrated that acinar cells also posses endogenous NOS activity14). Furthermore, the salivation induced by cholinergic agonists or that produced by the chorda stimulation was decreased by inhibition of NO synthesis by NG-nitro-L-arginine methyl ester (L-NAME) in the rat submandibular gland11, 15).
Nevertheless, the role of NO in the regulation of sodium transporters and AQP channels in the salivary gland has not been established. Therefore, the present study was aimed to explore the role of NO in the regulation of sodium transporters and AQP channels in the salivary gland. Rats were inhibited of endogenous generation of NO by treatment with L-NAME, and the expression of sodium transporters and water channels was determined in the submandibular gland.
Materials and Methods
1. Animals
Male Sprague-Dawley rats (200-250 g) were used. The experimental group was treated with L-NAME (100 mg/L drinking water) for 1 week. The control group was supplied with normal tap water. The experimental procedure conformed to the Institutional Guidelines for Experimental Animal Care and Use.
2. Western blot analysis
The submandibular glands were rapidly isolated under ketamine anesthesia. They were rapidly frozen and kept at -70℃ until analyzed. For protein preparation, they were thawed and homogenized with Polytron homogenizer at 3,000 rpm in a solution containing sucrose (250 mmol/L), EDTA (1 mmol/L), phenylmethylsulfonyl fluoride (0.1 mmol/L), and potassium phosphate buffer (20 mmol/L), at pH 7.6. The homogenate was centrifuged at 1,000 g for 15 min to remove the whole cells, nuclei, and mitochondria. The supernatant was then ultracentrifuged at 100,000×g for 1 h to produce a pellet containing membrane fractions enriched for both plasma membranes and intracellular vesicles. The pellet was resuspended in homogenizing solution for protein blotting.
Protein samples were loaded and electrophoretically size-separated with a discontinuous system consisting of 8-12.5% polyacrylamide resolving gel and 5% polyacrylamide stacking gel. The proteins were then transferred to a nitrocellulose membrane at 40 V for 3 h. The membrane was washed in Tris-based saline buffer (pH 7.4) containing 0.1% Tween-20 (TBST), blocked with 5% nonfat milk in TBST for 1 h, and incubated with antibodies in 2% nonfat milk/TBST for 1 h at room temperature. The antibodies used were polyclonal anti-rabbit α-1 and β-1 subunits of Na+,K+-ATPase (1:2,500, Upstate Biotechnology, Lake Placid, NY, USA), NKCC2 (1:1,000, Chemicon, Temecula, CA, USA), NHE1 (1:500, Alpha Diagnostic, San Antonio, TX, USA), α-subunit of ENaC (1:500, Alpha Diagnostic, San Antonio, TX, USA), AQP1 (1:1,000, Alomone, Jerusalem, Israel), AQP5 (1:1000, Alpha Diagnostic, San Antonio, TX, USA), endothelial NOS (eNOS) and neuronal NOS (nNOS) (1:750, Transduction; Lexington, KY, USA). β-Actin was used as internal control (1:1,000, Sigma, St. Louis, MO, USA). The membrane was then incubated with horseradish peroxidase-labeled goat anti-rabbit IgG (1:1,200) in 2% nonfat milk in TBST for 2 h. The bound secondary antibody was detected by enhanced chemiluminescence (Amersham, Buckinghamshire, England). The protein levels were determined, using the transmitter scanning videodensitometer (Biomed Instruments, Fullerton, CA, USA).
Results
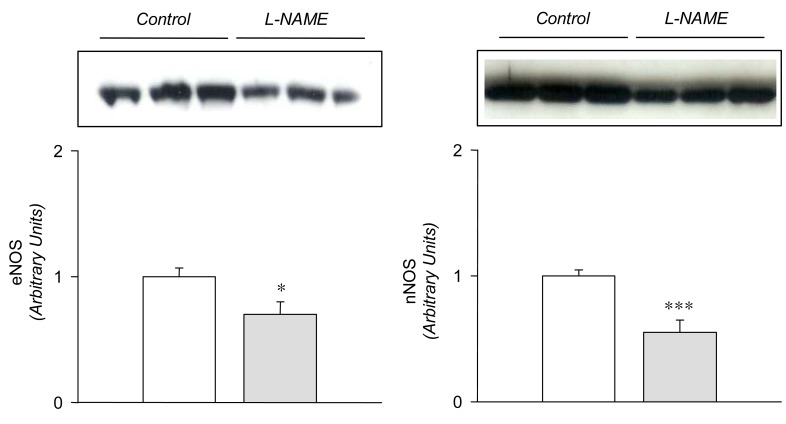
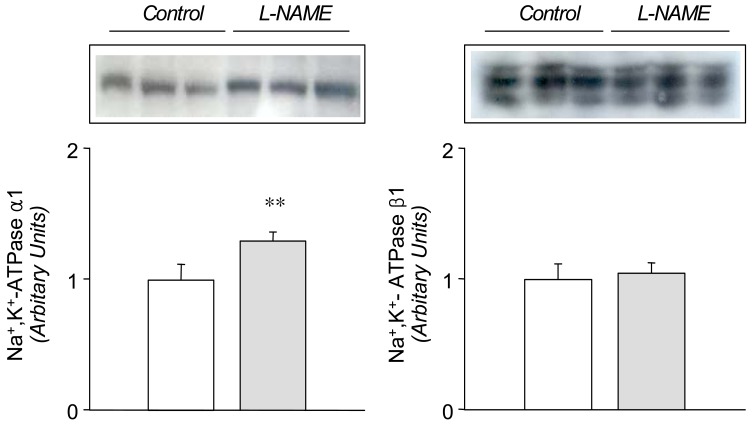
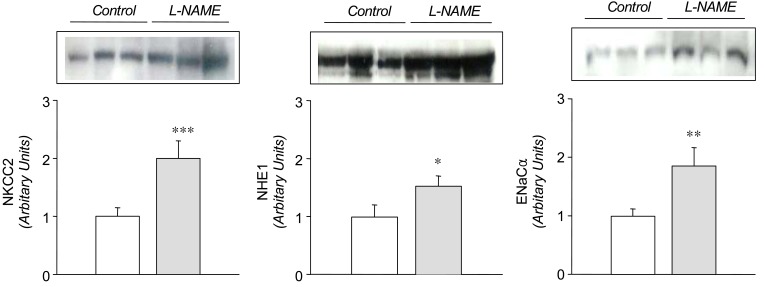
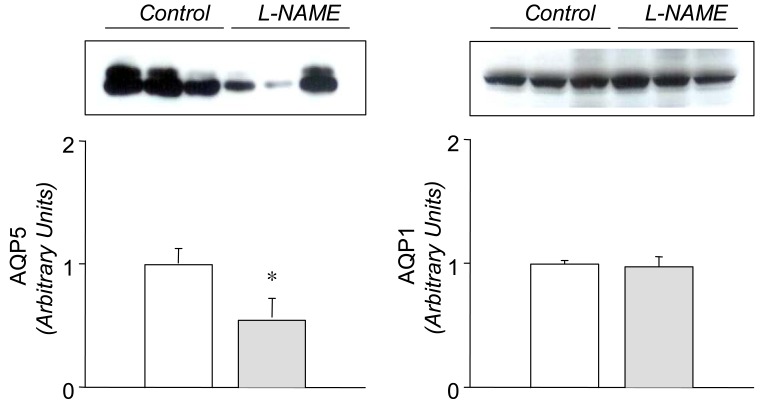
In the L-NAME-treated group, the systolic blood pressure measured indirectly by the tail cuff method in a conscious state was significantly higher in the experimental group than in the control (146±9 vs 121±6 mmHg, p<0.05, n=6 each). Accordingly, the expression of eNOS and nNOS in the submandibular gland was decreased significantly (Fig. 1). The amount of β-actin was not significantly altered (Fig. 2). On the contrary, the expression of α1-subunit of Na+,K+-ATPase was significantly increased, although that of β1-subunit remained unchanged (Fig. 3). The expression of NKCC2, NHE1, and α-subunit of ENaC was increased significantly (Fig. 4). The expression of AQP5 was significantly decreased, while that of AQP1 was not significantly altered (Fig. 5).
Discussion
NO has been implicated in mechanisms mediating nerve-evoked vasodilator and secretory responses in salivary glands. The salivation induced by cholinergic agonists or chorda stimulation was decreased by NO synthesis inhibition in the submandibular gland12, 13, 15). The formation of primary and secondary saliva involves transepithelial electrolyte and water movement. Therefore, it is plausible to hypothesize that NO plays a role in regulating the sodium transporters and AQP channels in the salivary gland.
Previous studies have shown that L-NAME treatment decreases the gene and protein expression of eNOS16, 17). The present study also demonstrated a decreased expression of both eNOS and nNOS in the submandibular gland following the treatment with L-NAME, which may have resulted in a decreased formation of NO. The down-regulation of NOS was associated with an increased expression of a1-subunit of Na+,K+-ATPase, NKCC2, NHE1, and a-subunit of ENaC, suggesting that these sodium transporters should be under tonic inhibitory influence of NO in the submandibular gland. The up-regulation of these transporters may then, at least in part, account for the decreased salivation following inhibition of NO synthesis in rat submandibular gland13, 15), possibly through an enhancement of the ductal reabsorption of sodium.
On the other hand, the expression of AQP5 was significantly decreased by the treatment with L-NAME. This finding suggests that AQP5 is under tonic excitatory influence of NO in the submandibular gland. A key role of AQP5 in saliva fluid secretion has been well known. Transgenic mice lacking AQP5 display reduced pilocarpine-stimulated saliva secretion (60%), in which the secreted saliva is more hypertonic and viscous18). In Sjøgren's syndrome, an abnormal distribution of AQP5 has been demonstrated in the acini, which is likely to contribute to the deficiency of fluid secretion19). A down-regulation of AQP5 may result in a decrease of water flux in the acinar cell and hence the formation of primary saliva. In this context, the L-NAME-induced reduction of secretory responses12, 13, 15) may be attributed in part to the down-regulation of AQP5. The expression of AQP1 has been demonstrated to be continuously distributed to microvasculature during embryonic and postnatal development in the rat submandibular gland20, 21). However, the expression of AQP1 proteins was not significantly altered by the L-NAME-treatment. In transgenic mice lacking AQP1, pilocarpine-induced salivary secretion showed no defects in volume or composition of saliva18, 22). It is unlikely that AQP1 should play a major role in salivary secretion.
Finally, it is worth to note that the systolic blood pressure was significantly increased, indicating that the animals were in a state of systemic withdrawal of tonic influences of endogenous NO. Furthermore, the alteration of sodium transporters and water channels cannot be attributed to the increased blood pressure, perse, since an attenuated salivation has not been documented in other models of hypertension.
In summary, these results indicate that sodium transporters and water channels are under tonic regulatory influence of NO in the submandibular gland. An altered NO synthesis may affect the expression of these transporters and channels, and hence the formation and secretion of saliva. Further studies will be needed to directly measure the sodium and water flux in the ductal epithelia to determine the actual contribution of each transporter.
 XML Download
XML Download