

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Protein kinase C (PKC) is a family of protein kinases that specifically phospholylate serine/threonine residues. The family includes at least 11 isoforms (α, βI, βII, γ, δ, ε, ζ, η, θ, λ, and µ) in mammalian tissue. These isoforms are divided into three subgroups based on their structure and mode of activation. The first group, the classical or conventional PKCs (cPKCs), including the isoenzymes α, βI, βII and γ, are dependent on activation through diacylglycerol (DAG) and Ca2+. The second group, the new or novel PKCs (nPKCs), including δ, ε, η θ and µ, are activated by DAG. The third group, the atypical PKCs (aPKCs), including ζ and λ, are not activated by DAG or Ca2+1-3).
PKC plays a central role in intracellular signal transduction pathways for hormones, neurotransmitters and growth factors, and significantly contributes to the control of various renal functions, including cellular proliferation, differentiation, exocytosis, and ion and water transport3-5).
In cultured cells, PKC inhibits activation of Na+,K+-ATPase4,6) and activates the Na+/H+-exchanger and Na+/HCO3- cotransporter7,8). According to the findings, PKC may be involved in the modulation of intracellular transporters.
There are several studies showing that various PKC isoforms that are expressed in the rat kidney. Kosaka et al.9) and Ono et al.10) showed the PKC isoenzymes α, β, and ζ; Wetsel et al.11), α, βII, δ, ε, and ζ; Caterina et al.12) and Aristimuno and Good13), α, β, δ, ε, and ζ; Ostlund et al.14) and Serlachius et al.15), α, δ, ε, and λ; and Pfaff et al.16), α, βI, and βII. Although most of these studies identified the PKC isoenzymes using molecular biologic approaches, little is known about their localization along the nephron.
Hashimoto et al.17) and Hirataet al.18) detected PKC in brain tissue and Puceat et al.19) and Rybin et al.20) detected it in heart tissue. Serlachius et al.15) suggested a distinct and differential expression and distribution of PKC isoenzymes depending on embryonal development in the kidney. Moreover, they reported that inhibition of PKC activation enhances apoptosis and induces impairment of nephron formation. These findings support that PKC plays a role in growth and differentiation in development21-25).
To identify the function of PKC in the kidney, we studied the differential expression and localization of the PKC isoenzymes α, βI, βII and γ in the developing rat kidney using immunohistochemistry.
Materials and Methods
1. Animals and Preservation of Kidneys
Male Sprague Dawley rats weighing approximately 250 to 300 g were used in all experiments. Prenatal kidneys were obtained from 16-, 18- and 20-day-old fetuses. Postnatal kidneys were obtained from 1-, 3-, 7-, 14- and 21-day-old pups and adult. The animals were anesthetized with an intraperitoneal injection of urethane (16.5%) and perfused with periodate-lysine-paraformaldehyde (PLP) solution for 3-5 minutes through the abdominal aorta. Kidneys were removed, and cut into 2-mm-thick slices, including the renal papilla. Slices were then immersed in PLP solution for 6-12 hours at 4℃. Tissues were embedded in wax or EPON 812. For immunohistochemistry using a pre-embedding method, PLP-fixed tissues were cut on a vibratome (Lancer Vibratomes Series 10 00; Technical Products International, St. Louis, MO) to a thickness of 50 µm.
2. Immunohistochemistry
1) Immunostaining of wax sections
The 50-µm-thick wax sections were dewaxed in xylene and hydrated through an ethanol series and washed for 10 minutes. Sections were incubated with 1.4% methanolic H2O2 for 30 minutes and with 0.5% Triton X-100 (0.01 M PBS, pH 7.4) for 15 minutes. After rinsing three times in PBS, sections were incubated for 1 hour in PBS containing 10% normal goat serum (Vector Laboratories, Burlington, CA, USA). Sections were immunostained with rabbit polyclonal IgGs (Santa Cruz technology, CA, USA) against PKCs α, βI, βII and γ as primary antibodies, using a Vectastain ABC kit (Vector Laboratories) according to the manufacturer's instructions. The sections were incubated overnight in PBS solutions containing antibodies diluted 1:2000 for PKCα, 1:2000 for PKCβI, 1:1500 for PKCβII and 1:5000 for PKCγ. Tissue sections were washed three times for 10 minutes and then incubated for 2 hours at room temperature with PBS containing biotin-conjugated goat anti-rabbit IgG (Vector Laboratories), diluted 1:500, as the secondary antibody. Avidin-biotin-peroxidase complex (Vector Laboratories) diluted 1:100 was used as the tertiary reagent. After the sections were washed three times for 10 minutes with 0.05 M Tris-HCl buffer (pH 7.6), the 0.05% diaminobenzidine and 0.0033% H2O2 were used as chromogen. After the immunostaining, sections were counterstained with hematoxylin.
2) Immunostaining of 1-mm-thick EPON sections
To identify the immunoreactivity of PKC in intercalated cells in adult rat kidney, 1 mm semi-thin sections, embedded in EPON-812, were cut into slices displaying cortex, outer medulla and inner medulla. The EPON was removed using saturated sodium hydroxide. Antibodies against PKCα, βI, βII and γ were used. H+-ATPase (1:2,000) were used on adjacent sections. Antibodies against aquaporin-1 (AQP-1; 1:2,000) were used to differentiate descending thin limbs of Henle from proximal convoluted tubules. Immunostaining was performed with avidin-biotin-peroxidase complex (ABC), and then the sections were examined after staining with the blue-gray-colored Vector SG (Vector Laboratories).
Results
1. Immunohistochemistry
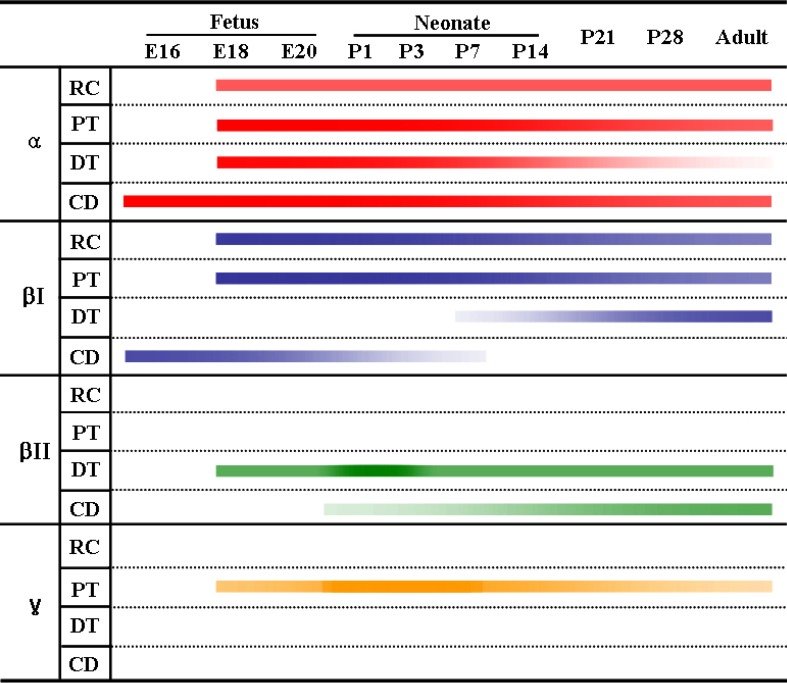
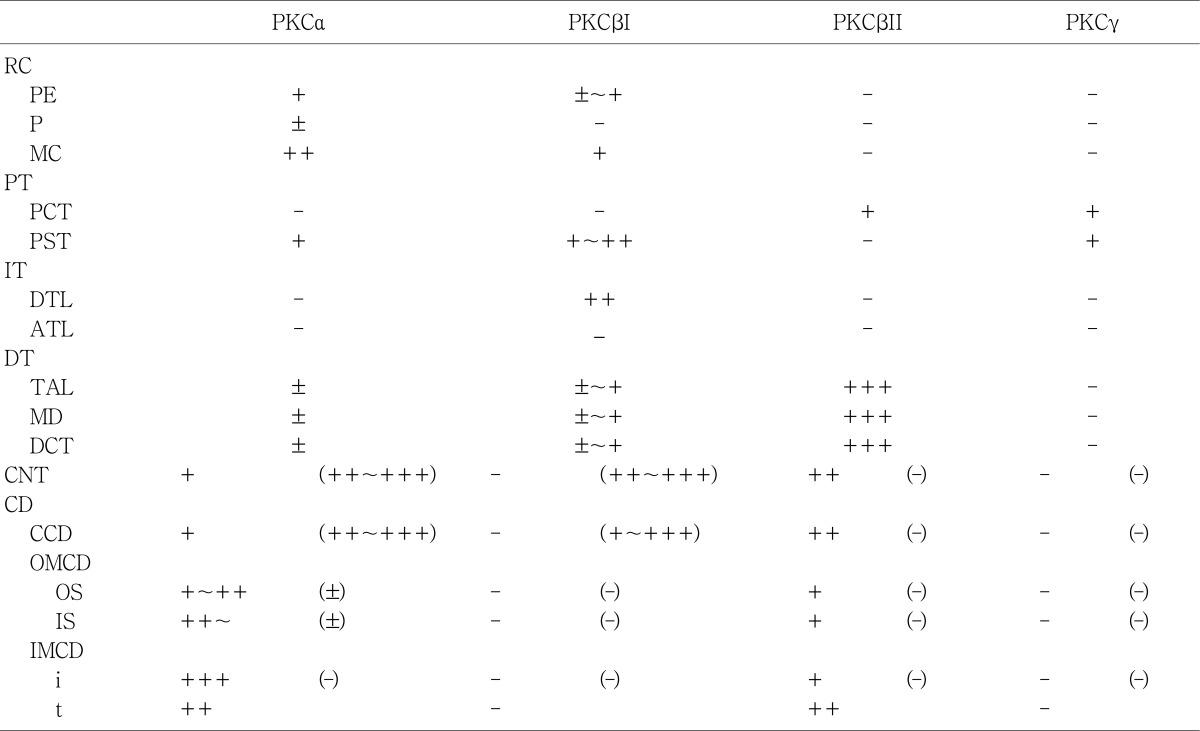
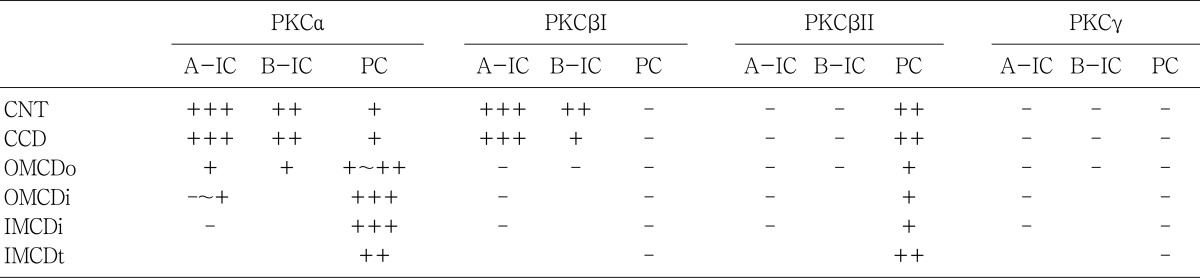
The PKC isoenzymes α, βI and βII, but not γ, were expressed in the adult rat kidney in the tubules, and the distribution in the tubules was variable (Table 1, 2). PKC isoenzymes α, βI, βII and γ were expressed in the developing kidney, and distinct and differential expression patterns were shown during development.
2. PKCα
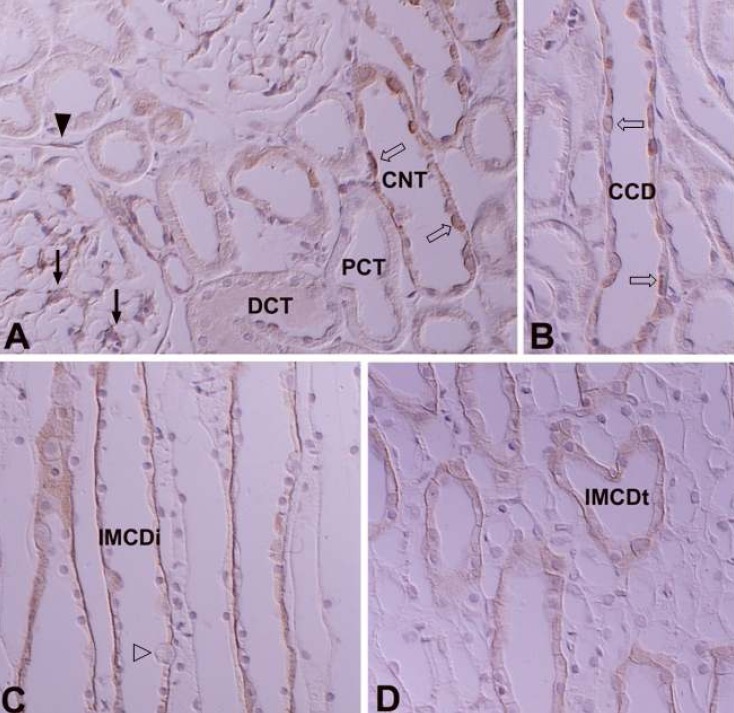
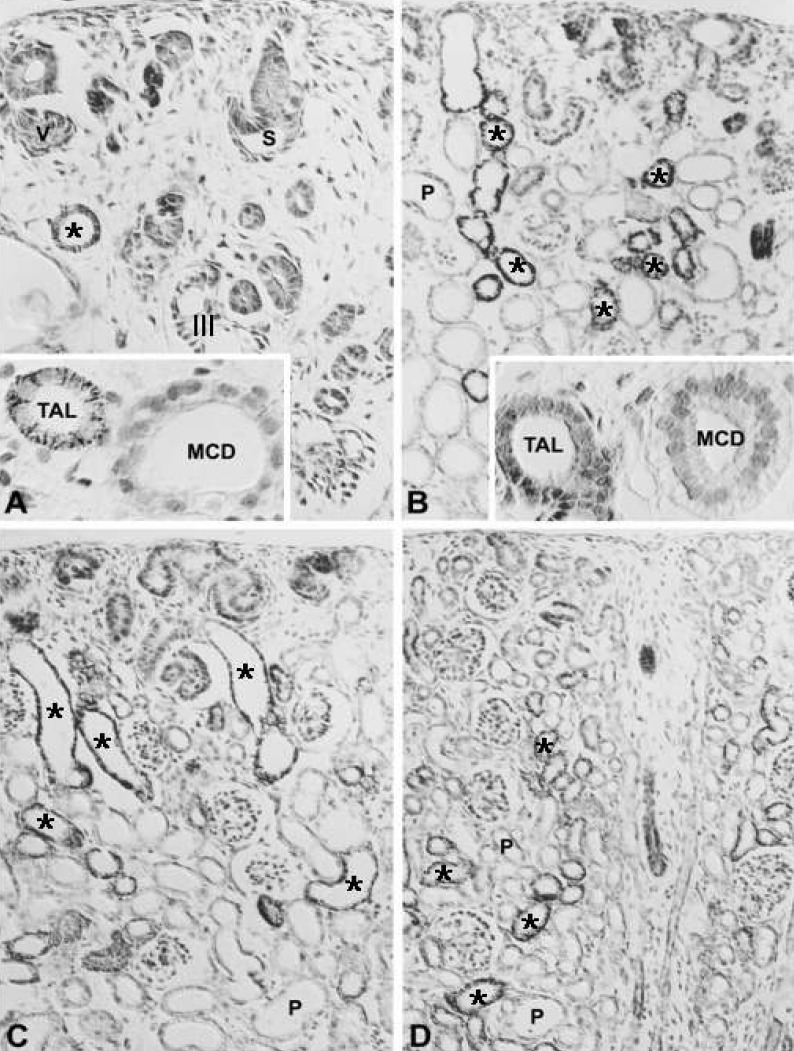
In the adult kidney, PKCα immunostaining was generally observed in the entire tubule, but was most strongly observed in the connecting tubules and the cortical collecting ducts. On staining with H+-ATPase, PKCα-positive cells were strongly evident in the intercalated cells. Type A intercalated cells were stained in the supranuclear portion of the cytoplasm and type B intercalated cells were stained throughout the entire cytoplasm. The connecting tubule cells and principal cells were stained weakly at the basolateral plasma membrane. In the medullary collecting ducts, the intercalated cells were PKCα negative and the principal cells were PKCα positive at the basolateral plasma membrane. The inner stripe of the outer medulla and the initial part of the inner medulla showed strong immunoreactivity. In the renal corpuscle, mesangial cells showed moderate immunoreactivity, and we observed weak immunoreactivity in parietal epithelial cells and podocytes. In the proximal tubule, the convoluted part showed weak immunoreactivity on the microvilli and faint immunoreactivity was observed in the straight portion. The cytoplasm of distal tubule cells showed faint immunoreactivity. We did not observe any PKCα immunoreactivity in the descending or ascending limbs of the Loop of Henle (Fig. 1; Table 1, 2).
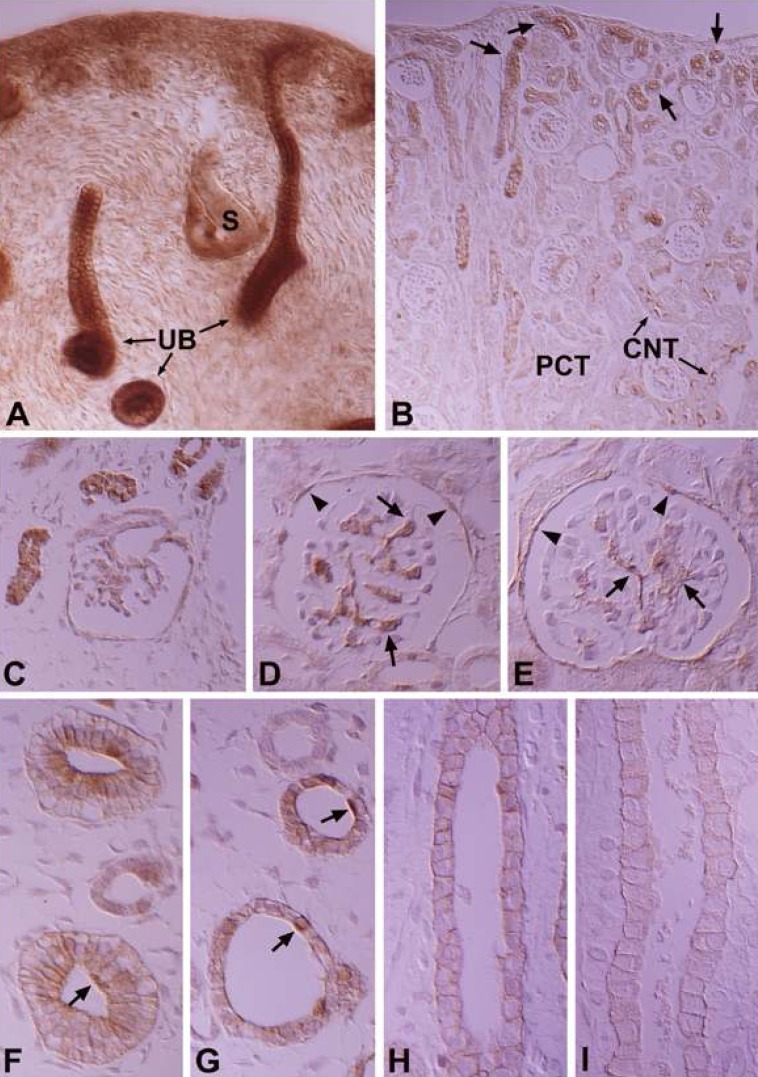
In the developing kidney, PKCα appeared in the ureteric bud at 16 days of gestation, but there was no staining of the renal vesicle and S-shaped body (Fig. 2A). The PKCα immunoreactivity of the collecting tubule gradually decreased during development and showed a mature pattern from 14 days after birth (Fig. 2F-I). PKCα appeared strongly in the mesangial and parietal cells of the developing renal corpuscle in stage III, proximal anlage and distal anlage of the 18-day-old pups, whereas immunoreactivity for PKCα gradually decreased in mature proximal convoluted and distal convoluted tubules (Fig. 2A-E). In the intercalated cells, immunoreactivity was shown in the connecting and collecting tubules of 18-day-old pups.
3. PKCβI
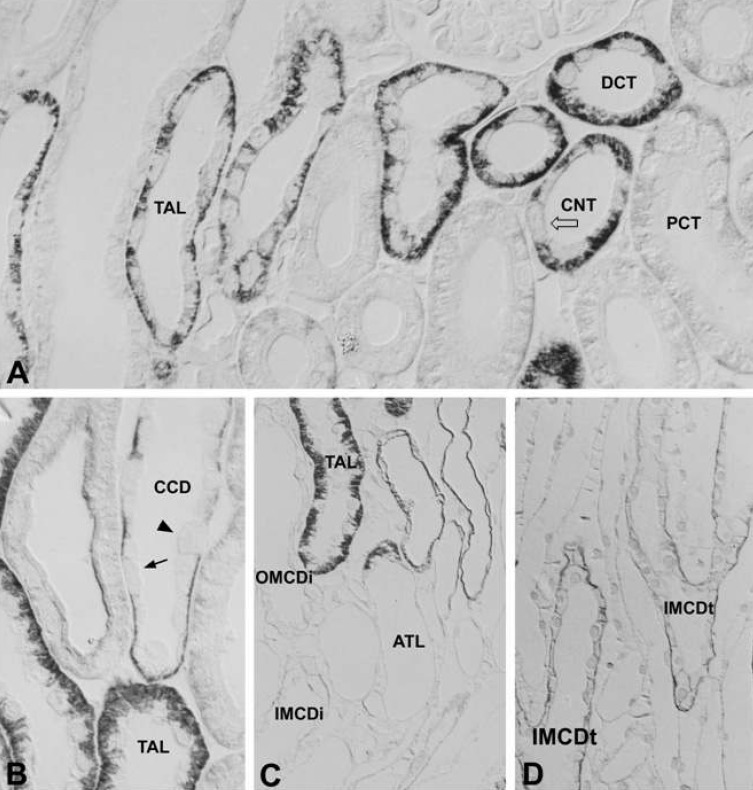
In the adult kidney, there was strong positive PKCβI staining in the connecting segment and intercalated cells of the cortical collecting duct. Similar to PKCα, type A intercalated cells were positive in the supranuclear area and type B cells were positive throughout the entire cytoplasm. PKCβI staining was negative in principal cells. In the proximal tubule, the convoluted part was negative and the straight portion was moderately positive. In the renal corpuscle, the mesangial cells were weakly positive, parietal cells faintly positive, and the podocytes negative. The thick ascending limb of the Loop of Henle and the distal convoluted tubule were also negative. We did not observe immunoreactivity in the outer and inner medullary collecting tubules. However, we did detect moderately positive staining in the inner medulla, at the apical plasma membrane of the descending thin limb (Fig. 3; Table 1, 2).
In the developing kidney, PKCβI immunoreactivity appeared from 16 days of gestation and was strongly positive in the ureteric bud (Fig. 4A). The immunoreactivity of the collecting tubule was strong in the fetus, but decreased markedly after birth, and the principal cells were negative 3 days after birth (Fig. 4E-G). In the renal corpuscle, mesangial cells, parietal cells, and the proximal anlage were strongly positive in 18-day-old pups (Fig. 4B, C). The renal vesicles and S-shaped bodies were negative (Fig. 4A). During development, the straight portion of the proximal anlage remained moderately immunopositive and the immunoreactivity in the convoluted portion disappeared after birth, being negative from 14 days after birth. In the distal nephron, the distal anlage was negative, with immunoreactivity only becoming positive from 21 days after birth. PKCβI immunoreactivity in the intercalated cells showed a pattern similar to PKCα in appearance and distribution.
4. PKCβII
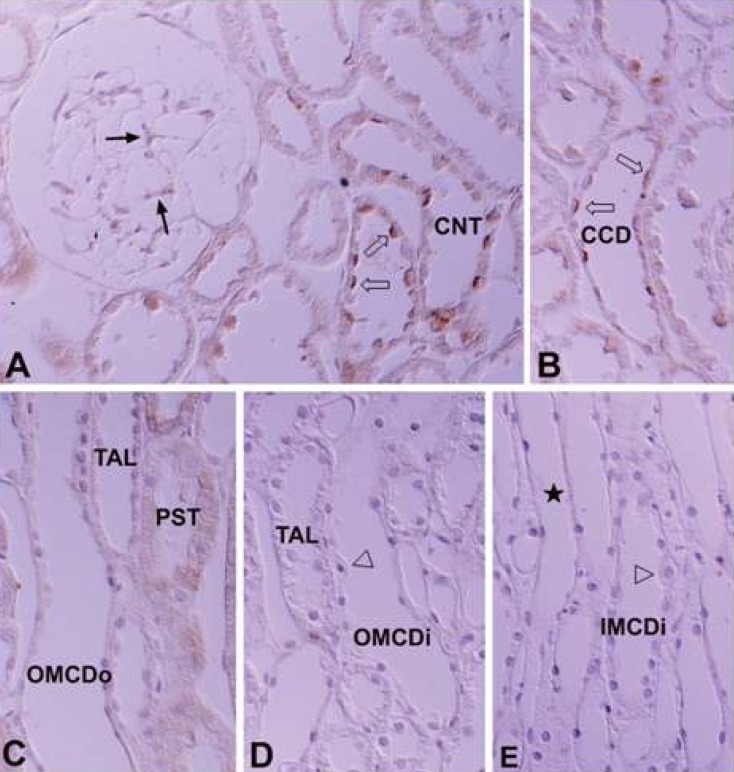
In the adult kidney, immunoreactivity for PKCβII was strongly positive in the thick ascending limb of the Loop of Henle, the macula densa, the distal convoluted tubule, and the basolateral membrane of the connecting tubule. In the collecting tubule, the basolateral membrane of the principal cells was moderately positive, but no immunoreactivity was seen in the intercalated cells of the connecting and collecting tubules (Fig. 5B-D). There was weak basolateral labeling in the proximal convoluted tubule. There was no immunoreactivity in the renal corpuscle or intermediate tubule (Fig 5A; Table 1, 2).
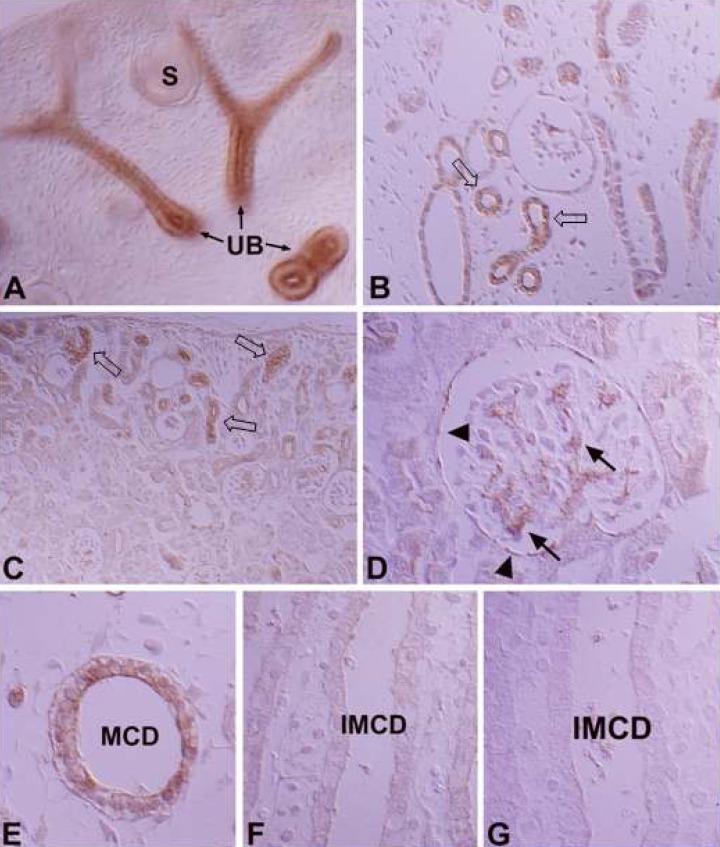
In the developing kidney, PKCβII immunoreactivity appeared in the basolateral membrane of the distal anlage at 18 days of gestation. Immunoreactivity increased markedly in the distal tubule, including the thick ascending limb of the Loop of Henle, the distal convoluted tubule and the connecting tubule from 1 day after birth. PKCβII immunoreactivity gradually decreased from 7 days after birth and had a similar pattern to the adult rat from 21 days. The intercalated cells in the connecting and collecting tubules were negative for PKCβII immunostaining. The principal cells were negative during the initial stages of development, but immunoreactivity gradually increased after birth and showed a similar pattern to adult rats from 14 days after birth (Fig. 6).
5. PKCγ
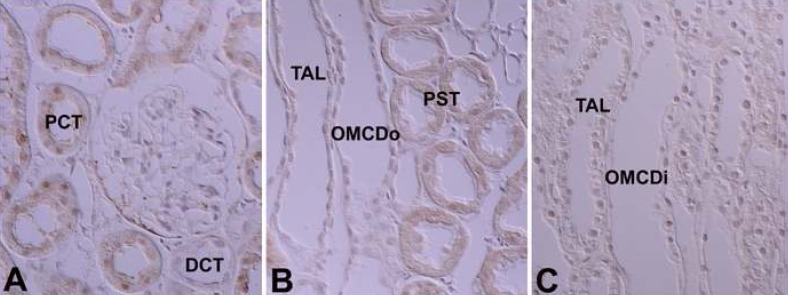
There was very weak immunoreactivity for PKCγ only in the proximal tubule, whereas no immunoreactivity in other uriniferous tubules (Fig. 7; Table 1, 2). In the developing kidney, PKCγ immunoreactivity was strong in the proximal tubule. Immunoreactivity appeared in the proximal anlage at 18 days of gestation. There was strong positive staining in the entire proximal tubule at 1, 3, and 20 days after birth. Subsequently, immunoreactivity decreased and had a similar pattern to adult rats from day 21 after birth (Fig. 8).
Discussion
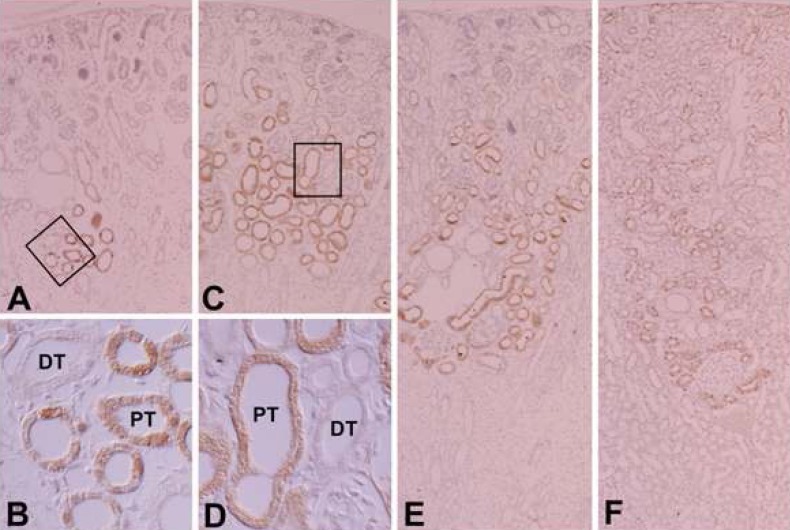
PKC plays a central role in intracellular signal transduction. The various PKC isoforms are expressed in the rat kidney with distinct and differential expression patterns (Fig. 9). As a member of the cPKC group, PKCα expression was predominant in the adult kidney. PKCβ was localized in the tubules. PKCγ is known to be detected in the central nervous system10). Our study, using immunohistochemistry and immunoblotting, demonstrates that the expression of PKCα, βI and βII, but not PKCγ, is evident in in the tubules of the rat kidney and PKCα, βI, βII, and γ are expressed in the developing kidney.
Wetsel et al.11), Caterina et al.12), Dong et al.22), Pfaff et al.26), Saxena et al.27), Ostlund et al.14), Aristimuno and Good13), and Serlachius et al.15) have reported the expression of PKCα in the kidney. In our study, the expression of PKCα was detected in the cortex, outer stripe of the outer medulla, inner stripe of the outer medulla and, using immunoblotting, in cytosolic and membrane fractions from the inner medulla. Using immunohistochemistry, Dong et al.28) and Fukuzaki et al.29) reported PKCα expression in the renal corpuscle, proximal straight tubule and collecting duct of the inner medulla of rat and human kidneys. However, our study demonstrates that PKCα staining was diffusely positive, with the exception of the intermediate tubule. Especially, strong positive staining was observed in the connecting tubule, intercalated cells of the cortical collecting tubule, mesangial cells of the renal corpuscle, outer and inner stripes of the outer medulla, and the principal cells in the collecting duct of the inner medulla.
Wetsel et al.11), Ostlundetal.14) and Aristimuno and Good13) reported the expression of PKCβI in the kidney, but not PKCβII. Caterina et al.12) identified PKCβ in renal corpuscles using immunohistochemistry. Our study demonstrates the expression of PKCβI and βII. On immunoblotting, PKCβI was faintly detected in the cortex and outer stripe of the outer medulla in the cytosolic fractions. Expression was generally weakly detected in the membrane fraction. Using immunohistochemistry, PKCβI immunoreactivity was detected in the connecting segment, cortical collecting tubules, proximal straight tubules, mesangial cells of the renal corpuscle and the parietal epithelium. The connecting tubules and intercalated cells of the cortical collecting tubules were strongly positive. The apical plasma membrane of the descending thin limb of Henle was positive. These results were consistent with the immunoblotting findings.
A PKCβII band was observed in the membrane fractions. On immunohistochemistry, PKCβII was expressed in the proximal convoluted tubules, distal convoluted tubules, and the basolateral plasma membrane of the connecting and collecting tubules. The distal convoluted tubules, including the thick ascending limb of the Loop of Henle, and the connecting tubules were strongly positive. The intercalated cells in the distal nephron showed distinct and different expression patterns for PKC isoenzymes. In the connecting segment and cortical collecting tubules, the intercalated A cells play a role in H+ secretion and the intercalated B cells are involved in HCO3- secretion. In the medullary collecting tubules, intercalated cellsplayarole in H+ secretion30). Our study shows that the type A intercalated cells in the connecting and cortical collecting duc tare strongly positive for PKCα and β in their supranuclear cytoplasm. Type B intercalated cells were moderately positive in their cytoplasm and basolateral plasma membranes. Those findings were consistent with the results of mouse kidneys31) and the location of the H+-ATPase32), so we suggest that PKCα and βI may contribute to secretion of protons. However, PKCα, and βI were not expressed in the intercalated cells in the medullary collecting ducts, so we speculate that different control mechanisms exist between intercalated cells in the connecting segment and cortical and medullary collecting tubules.
Several studies have reported that PKCγ is not detected in the adult rat kidney11,13,14,22). Recently, studies have shown that PKC plays a role in growth and differentiation during development21-24). Serlachius et al.15) suggested that there is distinctand differential expression and distribution of PKC isoenzymes depending on embryonal kidney development. We also observed distinctand differential expression and distribution of PKC isoenzymes depending on kidney development. PKCα, βI, and βII were expressed and, interestingly, PKCγ was detected temporarily in the developing kidney. Positive staining for PKCα and βI appeared in the ureteric bud at 16 days of gestation and was strongly positive before birth, and then gradually decreased after birth. Therefore, we suggest that PKCα and βI play a role in differentiation of the collecting tubule. PKCα expression was positive in the proximal and distal anlage of pups up to 7 days of age. Therefore, we believe that PKCα expression is correlated with differentiation of the proximal and distal tubules. Moreover, PKCβI expression is correlated with differentiation of the proximal tubule because it was temporarily expressed in the proximal anlage in early stages of development. PKCβII appeared in the basolateral membrane of the distal and connecting tubules at 18 days of gestation, and gradually increased during development. Therefore, we suggest that PKCβII expression is not correlated with growth and differentiation. PKCγ immunoreactivity appeared and was highly expressed in the proximal anlage of the 18-day-old fetus, then decreased markedly, and disappeared soon after birth. Therefore, PKCγ appears to be correlated with differentiation of the proximal tubule.
In summary, our study demonstrates that the classical PKC isoforms, PKCα, βI and βII, but not PKCγ, are expressed in the tubules of adult rat kidneys, and PKCα, βI, βII, and g are expressed in the developing kidney, and there are distinct and differential expression patterns for the isoforms according to location and stage of development.
 XML Download
XML Download