

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Obesity is associated with various metabolic conditions, including hypertension, diabetes, dyslipidemia, and atherosclerosis, and is considered to be a growing public health concern [1]. One of the main causes of obesity is leptin resistance, which is associated with hypothalamic inflammation [2]. It is well-known that a high-fat (HF) diet induces hypothalamic inflammation in rodents and that the intracerebroventricular administration of saturated fatty acids elevates the expression of inflammatory cytokine in the hypothalamus [3]. Furthermore, the suppression of hypothalamic inflammation with a specific inhibitor reduces appetite and body weight in experimental animals treated with an HF diet [4].
Toll-like receptors (TLRs) initiate host defense mechanisms in response to microbial products, DNA, and RNA [56]. Activated TLRs trigger an intracellular signaling pathway to produce inflammatory cytokines, such as interleukin-1β, tumor necrosis factor-α, and interleukin-6 [78]. Moreover, TLR4 is involved in the pathogenesis of obesity [4] and provides a link between fat and inflammation [9]. Intracerebroventricular treatment with TLR4 antiserum for 7 days reduces the expression of hypothalamic inflammatory cytokines and normalized leptin resistance [10]. Treatment with polyunsaturated fatty acids attenuates inflammation [11], suppresses HF diet-induced activation of TLR4, and prevents gains in body weight [12]. Thus, we hypothesized that suppression of TLR4 in the hypothalamus would promote weight reduction.
Park et al [13] reported that udenafil, a phosphodiesterase 5 (PDE5) inhibitor prescribed for erectile dysfunction, suppressed TLR4 in human airway epithelial cells. A 14-year prospective study identified obesity as a risk factor for erectile dysfunction [14] and a 12-week HF diet decreased erectile function in rats [15]. Moreover, weight reduction with bariatric surgery improved erectile dysfunction significantly in obese men [16]. Recently, Moon et al [17] reported that daily administration of udenafil (75 mg/d) for 6 months improved erectile dysfunction in a randomized placebo-controlled clinical trial. Thus, it is possible that the suppressive effect of udenafil on TLR4 induced body weight reduction in obese conditions, and the reduction in body weight caused by udenafil, would be beneficial for improving erectile dysfunction, especially in obese men receiving long-term, daily treatment. However, the anti-obesity effects of udenafil have not yet been reported.
The purposes of the current investigation were to determine whether udenafil suppressed the hypothalamic expression of TLR4 mRNA and reduced body weight in HF-fed mice.
Go to : 
MATERIALS AND METHODS
1. Animal care
Male C57BL/6N mice (body weights: 18∼20 g) and Sprague-Dawley rats (body weights: 280∼300 g) were purchased from Jung-Ang Experimental Animals (Seoul, Korea). The animals were housed in individual cages with a 12-hour light/dark cycle (7:00 a.m. to 7:00 p.m.).
2. Ethics statement
Animals were cared for in accordance with the Guide to the Care and Use of Experimental Animals provided by the Yeungnam University Medical Center. The experimental protocol for this study was approved by the Ethics Committee of Yeungnam University.
3. Experimental design
1) Experiment 1 (direct injection)
To evaluate whether the expression of TLR4 was suppressed directly in the hypothalamus by udenafil (Zydena®; Dong-A Pharmaceutical Company, Seoul, Korea), we analyzed TLR4 mRNA expression 1 hour after intracerebroventricular injection. Udenafil (500 µg in 10 µL) or saline were infused over 5 minutes into the lateral ventricle of rats as described previously [18]. Briefly, a brain infusion cannula was placed into the lateral ventricle using a stereotaxic instrument under anesthesia with 8 mg/kg xylazine hydrochloride and 90 mg/kg ketamine. The coordinates were 1.3 mm posterior to the bregma, 1.9 mm lateral to the midsagittal suture, and a depth of 4.0 mm. The rat hypothalamus was excised 1 hour after the infusion and TLR4 mRNA expression was analyzed.
2) Experiment 2 (2-day treatment)
To evaluate udenafil-induced suppression of TLR4 mRNA, we used 24 control diet mice and divided them randomly and evenly into 4 groups. Udenafil was administered orally using gavage feeding needles at dosages of 0, 12, 120, or 600 µg/d for 2 consecutive days [18]. The dosages were determined according to a previous pharmacokinetic study for udenafil that showed significant changes at a dosage of 20 mg/kg/d for rats, which is analogous to ∼600 µg/d for mice [19]. Then, the hypothalamus was excised and mRNA expression of TLR4, PDE5, myeloid differentiation primary response gene 88 (Myd88), and nuclear factor-κB (NF-κB) were analyzed using real-time polymerase chain reaction (PCR).
3) Experiment 3 (9-day treatment)
To evaluate whether udenafil decreased body weight, we administered udenafil (500 µg/d, orally, once per day) or saline for 9 days to control- and HF-fed mice. Fifty-six mice were divided randomly into control and HF diet groups. The diets used in this study were HF with 60% of the calories from fat (HFD60%cal) and AIN-93G as the control diet. The HFD60%cal diet contained protein and carbohydrate (each at 20% of total calories) and was supplemented with vitamins (1%) and minerals (3.5%). The energy composition of the AIN-93G diet was 64%, 20%, and 16% carbohydrate, protein, and fat, respectively. The diets were purchased from Feedlab Korea (Seoul, Korea). These diets were fed to the mice for 8 weeks, a widely used protocol for the induction of obesity, leptin resistance, and various metabolic derangements in rodents [10]. Control- and HF-fed mice were divided into 4 groups; vehicle-, udenafil-, leptin-, and udenafil plus leptin- treated (n=7 in each group). Leptin (1 mg/kg/d, intraperitoneally) was administered in combination with udenafil or saline for 9 days to determine whether udenafil potentiated leptin sensitivity. Body weights and daily caloric intake were determined every morning. After 9 days of treatment, the mice were anesthetized with an intraperitoneal injection of tiletamine and zolazepam (25 mg/kg body weight, Zoletil®; Virbac, Carros, France). Blood was collected through the abdominal aorta with a heparin-coated syringe and then the plasma was separated by centrifugation at 4℃. The hypothalamus was excised and the expression of TLR4, pro-opiomelanocortin (POMC), and neuropeptide Y (NPY) mRNA was measured. Visceral fat mass (retroperitoneal and epididymal white adipose tissue) was excised and weighed.
4. Real-time polymerase chain reaction
Total RNA was obtained from the hypothalamus by using Trizol reagent (Bio Science Technology, Daejeon, Korea). cDNA was synthesized using 1 µg total RNA with oligo-(dT) primers and Prime RT Premix (Genet Bio, Nonsan, Korea). Real-time PCR was performed with duplicated samples in a LightCycler (Roche, Mannheim, Germany) as described previously [20]. The following primers were used: for mouse TLR4, (m)TLR4, 5′-AAC TAT TAC TGT CAA CGA CTT ACA GTT T-3′ (forward) and 5′-GCT TGG TCT TGA ATG AAG TCA-3′ (reverse); for rat TLR4, (r)TLR4, 5−-TCT ACG TGA AAA TCC CCA AGT-3′ (forward) and 5′-ATT CAT GGA GCC TAA TTC CC-3′ (reverse); for mouse NF-κB, 5′-CAG CTA CGG CGG CCT TCT-3′ (forward) and 5′-CTT TTG CGC TTC TCT TCA ATC C-3′ (reverse); for PDE5, 5′-TCA TCT TCT GTG GCT TGG G-3′ (forward) and 5′-ATG GTA AGA CAG GAC CTC TAA TGT-3′ (reverse); for MyD88, 5′-GCC CCA ATG ATA TCG AGT TT-3′ (forward) and 5′-TCT GTT CTA GTT GCC GGA TC-3′ (reverse); for GAPDH, 5′-AAC GGC ACA GTC AAG GCC-3′ (forward) and 5′-CGC TCC TGG AAG ATG GTG AT-3′ (reverse). These primers were designed by Primer Express software ver. 2.0 (Applied Biosystems, Foster City, CA, USA).
5. Data analysis
All results are expressed as mean±standard error. One-way analysis of variance was used to compare dose-dependent changes and the values among experimental groups. Post-hoc comparisons were performed using Duncan's test. Statistical comparisons between control and HF-fed mice were evaluated using Student's t-test. Statistical analyses were performed using SPSS software ver. 12 (SPSS Inc., Chicago, IL, USA). Values of p<0.05 were considered statistically significant.
Go to :
RESULTS
To determine whether udenafil suppressed the expression of TLR4 mRNA in the hypothalamus directly, we injected it into the lateral ventricle of rats. Udenafil significantly reduced the expression of hypothalamic TLR4 mRNA (p<0.05) (Fig. 1).
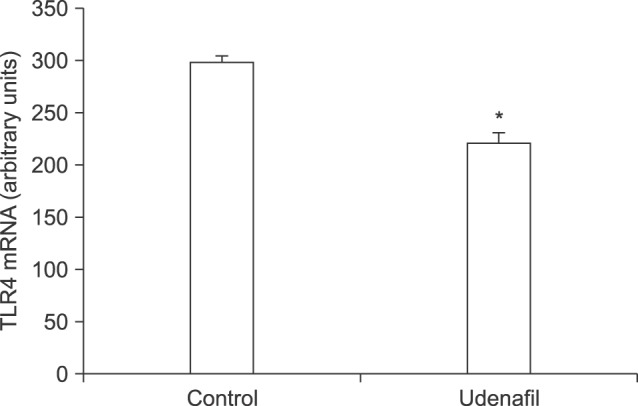 | Fig. 1The effect of udenafil on toll-like receptor 4 (TLR4) mRNA expression in the hypothalamus. Udenafil (500 µg) was dissolved in 10 µL saline. Udenafil or an equal volume of saline were injected into the lateral ventricle in rats. Data are expressed as mean±standard error of 3 experimental rats. *p<0.05 vs. control.
|
Oral treatment with udenafil for 2 days also suppressed the level of hypothalamic TLR4 mRNA in mice in a dose-dependent manner. This was accompanied by the suppression of PDE5, Myd88, and NF-κB mRNA in the hypothalamus (Fig. 2). PDE5 is the target molecule of udenafil, and Myd88 and NF-κB are signaling molecules for TLR4 activation.
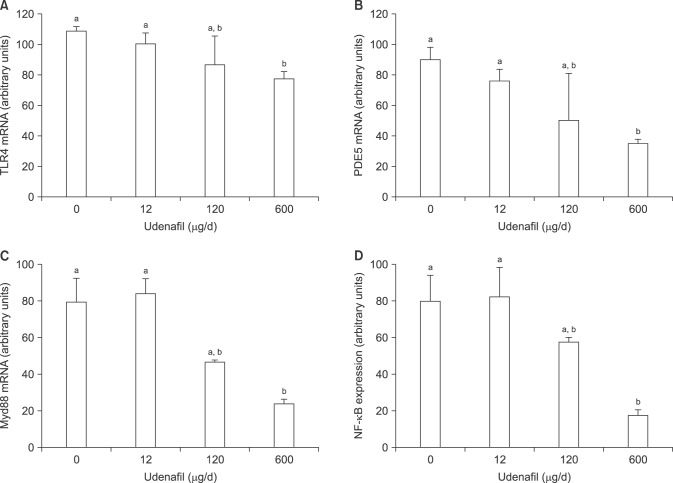 | Fig. 2The effects of different dosages (0, 12, 120, 600 µg) of udenafil on the expression of toll-like receptor 4 (TLR4) (A), phosphodiesterase 5 (PD5E) (B), myeloid differentiation primary response gene 88 (Myd88) (C), and nuclear factor-κB (NF-κB) (D) mRNA in the hypothalamus in mice. Udenafil was administered via gavage for 2 consecutive days. Data are expressed as mean±standard error of 6 mice in each experimental group. Values that do not share a common superscript letters are significantly different at p<0.05.
|
Obesity was induced by 8 weeks of HF feeding compared to control diet mice. The characteristics of HF-fed mice were increased body weight and visceral fat mass, and elevated plasma concentrations of leptin and total cholesterol compared to control diet mice (p<0.05) (Table 1).
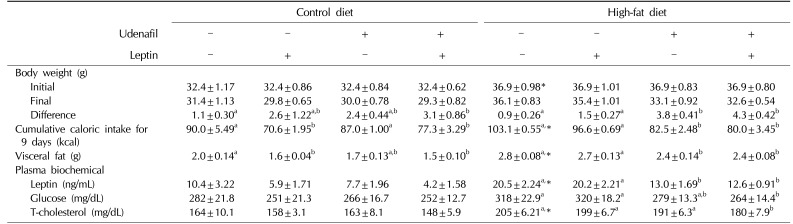
Table 1
Changes in physical and biochemical parameters following treatment of mice with udenafil and/or leptin for 9 days
![]()
Nine days' treatment with udenafil reduced body weight (p<0.01), visceral fat mass (p<0.05), cumulative caloric intake (p<0.01), and plasma leptin concentration (p<0.05) in HF-fed mice compared with vehicle-treated mice. Udenafil did not induce any statistically significant changes in body weight, visceral fat mass, or caloric intake in mice fed the control diet. Leptin treatment decreased cumulative caloric intake (p<0.05) and visceral fat mass (p<0.05) in mice fed the control diet; however, there were no specific effects in HF-fed mice. Combined treatment with udenafil and leptin decreased body weight, visceral fat mass, and caloric intake in both control diet and HF-fed mice compared to vehicle-treated mice. Moreover, the combined treatment suppressed the plasma concentrations of leptin (p<0.01), glucose (p<0.05), and total cholesterol (p<0.05) in HF-fed mice. However, the combined effects of leptin and udenafil were additive, rather than synergistic (Table 1).
After 9-day treatment with vehicle, udenafil, and/or leptin, the hypothalamus was excised and the expression of TLR4, POMC, and NPY mRNA was analyzed. Hypothalamic TLR4 mRNA expression appeared to be elevated in HF-fed mice compared with mice fed the control diet; however, there were no significant differences among the experimental groups because of the relatively large standard error found in the results. Although hypothalamic POMC mRNA levels showed a tendency to decrease in HF-fed mice, this difference was also not significant. Hypothalamic NPY mRNA expression was elevated in HF-diet mice compared with mice fed the control diet. This elevation was suppressed by udenafil and leptin treatment in both control diet- and HF-fed mice (Fig. 3).
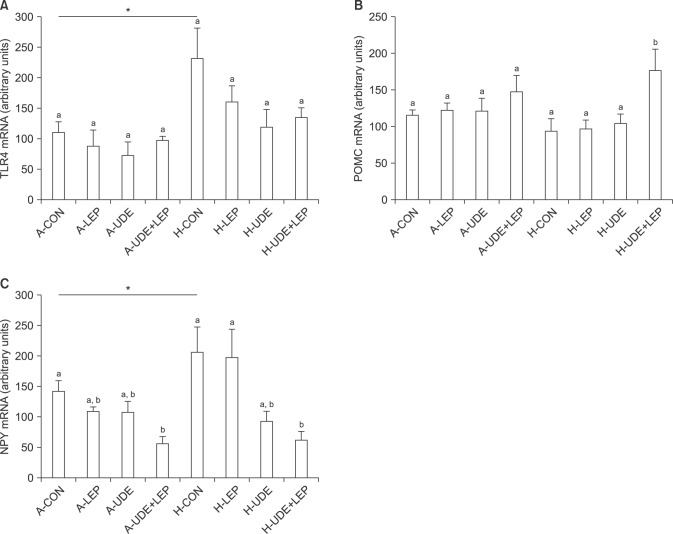 | Fig. 3Hypothalamic mRNA levels of toll-like receptor 4 (TLR4) (A), pro-opiomelanocortin (POMC) (B), and neuropeptide Y (NPY) (C) after treatment with udenafil and/or leptin for 9 days. The mRNA levels were analyzed using real-time polymerase chain reaction. A-: AIN-93G diet, H-: high-fat diet with 60% of the calories from fat, CON: control, LEP: leptin, UDE: udenafil. Data are expressed as mean±standard error of 7 mice in each experimental group. *p<0.05, vs. A-control. Values that do not share a common superscript letters are significantly different at p<0.05 in the control- or high-fat-fed mice.
|
Go to :
DISCUSSION
In the present study, we demonstrated that udenafil dose-dependently suppressed TLR4 mRNA expression in the hypothalamus and that the 9-day treatment of udenafil reduced body weight and visceral fat in HF-fed mice.
Hypothalamic inflammation is an important causative factor in the induction of obesity [2], and the reversal of hypothalamic inflammation restores body weight homeostasis [4]. During hypothalamic inflammation, TLR4 performs a critical role in initiating the inflammatory response [21]. Thus, we hypothesized that suppression of TLR4 would reduce body weight in obese conditions. In the present study, HF-fed mice showed elevated TLR4 mRNA expression in the hypothalamus, which was consistent with previous findings [312]. When saturated fatty acids or bacterial lipopolysaccharides bind to TLR4, intracellular signaling through Myd88 and NF-κB results in the secretion of several pro-inflammatory cytokines [7821]. We demonstrated that oral treatment with udenafil suppressed the hypothalamic expression of TLR4, which was accompanied by the suppression of Myd88 and NF-κB expression in a dose-dependent manner in mice.
It is well-known that obesity is frequently associated with erectile dysfunction. Anti-impotence drugs dilate penile blood vessels through elevation of the vasodilator, NO, via PDE5 inhibition [22]. Recently, it was reported that PDE5 inhibitors exerted anti-inflammatory effects [23]. Moreover, Varma et al [24] reported that chronic administration of tadalafil, a PDE5 inhibitor, improved metabolic parameters, such as fasting blood glucose and triglyceride levels, and reduced body weight; however, these changes were not statistically significant. No previous studies have evaluated how PDE5 inhibition was linked to the suppression of TLR4 or Myd88. The results of our current study confirmed the udenafil-induced suppression of TLR4, which was consistent with the results obtained by Park et al [13].
In the present study, we demonstrated that udenafil treatment for 9 days reduced body weight, visceral fat mass, and appetite in HF-fed mice, which was associated with a reduction in plasma leptin concentration. However, udenafil did not change body weight, caloric intake, or visceral fat mass significantly in mice fed the control diet. The present results are consistent with those obtained by Ayala et al [25], who showed that treatment with sildenafil for 12 weeks reduced the body weight of HF-fed mice by preventing diet-induced energy imbalance and insulin resistance. Taken together, the weight loss effect of udenafil in HF-fed mice seems to result from normalization of pathologic derangements induced by HF-feeding, as udenafil did not change body weight significantly in control- diet mice.
Treatment with 20 mg/kg/d udenafil for 4 weeks did not cause any specific toxicity to rats [19]. Our study used udenafil at approximately 13 mg/kg/d and also did not reveal any specific side effects of the chronic treatment. In a human study, Moon et al [17] administered 75 mg/kg/d of udenafil for 24 weeks and demonstrated an improvement in erectile function. Although the rate of adverse drug reactions was 17.9% in those patients, the reactions were mild and the patients recovered without additional treatment. Recent clinical trials have reported that daily treatment with PDE5 inhibitors allows a more spontaneous sexual life, shows excellent efficacy and safety, and requires a lower dosage than on-demand administration [2627]. Thus, the body weight reduction caused by udenafil in obese conditions demonstrated in the present study would be helpful for improving erectile dysfunction in obese men receiving the drug chronically.
Many mechanistic studies on the role of TLR4 in the induction of obesity have involved experiments in genemodified animals, in which the chemical compounds were required to affect TLR4 mRNA expression. We demonstrated hypothalamic udenafil-induced TLR4 suppression and observed an anti-obesity effect of udenafil in HF-fed obese mice. We expected that the combination of udenafil and leptin would exert a synergistic effect on body weight changes in HF-fed mice by improving leptin resistance; however, we only observed an additive effect.
In the hypothalamus, appetite is regulated mainly by a balance between the activities of orexigenic NPY-producing neurons and anorexigenic POMC-producing neurons [28]. Activation of POMC neurons by leptin suppresses appetite, whereas stimulation of NPY neurons increases food intake [29]. In the present study, HF-fed mice exhibited a relative suppression of POMC and elevation of NPY compared with mice fed the control diet, which was consistent with our previous study [18]. Interestingly, treatment with udenafil did not affect POMC mRNA expression, whereas udenafil suppressed NPY mRNA expression markedly in HF-fed mice. These effects may be associated with the suppression of caloric intake in HF-fed mice treated with udenafil, or the combined treatment of udenafil and leptin (Fig. 3). Although there are no specific reports supporting the transport of udenafil through the blood-brain barrier directly, Shim et al [30] suggested that this may occur. Their research demonstrated that daily treatment with udenafil appeared to improve cognition, depression and erectile function in patients with erectile dysfunction.
One limitation of this study was the absence of clarity around the mechanisms of the suppressive effect of udenafil on TLR4. Specifically, whether the inhibition occurred directly or secondarily via the inhibition of PDE5 is not known. Further studies are necessary to elucidate the mechanism of action of udenafil in the hypothalamus. Another limitation is that the factors that would affect the differences have not been controlled for statistically.
Go to :
CONCLUSIONS
The results of this study showed that udenafil suppressed TLR4 mRNA expression in the hypothalamus dose-dependently, and this was accompanied by the suppression of Myd88 and NF-κB expression. Nine-day treatment with udenafil reduced body weight and appetite in HF-fed mice, which was associated with the suppression of hypothalamic TLR4 and NPY levels. However, the effect of the combination of udenafil and leptin was additive rather than synergistic.
Go to :
 XML Download
XML Download