

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Peyronie's disease (PD) is a localized fibrotic process of the tunica albuginea (TA). The fibrotic plaque impedes the expansion of the TA during erection, which results in a variety of penile deformities and often pain during erection [12]. Although the etiology of PD is not fully delineated, an inflammatory process and subsequent aberrant wound healing following repeated trauma to the penis during intercourse are known to be involved in fibrotic processes [34]. Despite promising results with intralesional injection of collagenase clostridium histolyticum [5], surgical intervention is still the only curative treatment modality that corrects penile deformities [67]. Therefore, the identification of novel therapeutic target involved in complex fibrogenic process of PD is required.
Epigenetic modifications, such as histone acetylation/deacetylation and DNA methylation, comprise heritable alterations in the DNA itself without changes in the nucleotide sequence. Epigenetic modifications have been shown to play a crucial role in the pathogenesis of inflammatory disorders and fibrotic diseases [89]. Pharmacologic inhibition of histone deacetylase (HDAC) is known to decrease fibrotic responses in a variety of conditions [910]. Previous study demonstrated in skin fibroblasts from patients with systemic sclerosis that silencing HDAC7, a class II HDAC, specifically reduced excessive production of extracellular matrix, which was as effective as trichostatin A (TSA), a HDAC inhibitor. However, TSA also up-regulates the expression of profibrotic factors, such as connective tissue growth factor and intracellular adhesion molecule-1, whereas silencing of HDAC7 did not influence on the expression of these profibrotic molecules [11]. Therefore, inhibition of specific HDAC isoforms by use of RNA interference technology may be more advantageous than the use of nonspecific HDAC inhibitors.
In the present study, we examined the differential expression of HDAC isoforms in fibroblasts isolated from human PD plaque or normal TA. Next, we determined the effectiveness of the knockdown of HDAC7 on the transforming growth factor-β1 (TGF-β1)-induced profibrotic responses in primary fibroblasts derived from human PD plaque.
MATERIALS AND METHODS
1. Primary fibroblast culture
We obtained plaque tissues from two patients with PD (age, 48 and 52 years, respectively) or normal TA tissues from two control patients: 1 undergoing penoplasty for congenital curvature (age, 21 years) and 1 undergoing primary repair of TA as the result of penile fracture (age, 51 years). The tissue samples were used for primary fibroblast culture as previously described [1213]. Briefly, either plaque tissue or normal TA tissue was transferred into sterile vials containing Hank's balanced salt solution (GIBCO, Carlsbad, CA, USA) and was washed three times in phosphate-buffered saline (PBS). Biopsy tissue was minced into 1-mm2 segments and incubated in a shaker in 12.5 mL Dulbecco's modified Eagle Medium (DMEM) supplemented with 0.06% collagenase A (Sigma-Aldrich, St. Louis, MO, USA) for 1 hour. The cells and tissue fragments were collected by centrifugation (400 ×g, 5 minutes), washed in fresh culture medium, and placed in 100-mm cell culture dishes (Falcon-Becton Dickinson Labware, Franklin Lakes, NJ, USA) under standard cell culture conditions with DMEM supplemented with 10% fetal calf serum, penicillin (100 U/mL), and streptomycin (100 µg/mL). The dishes were incubated in a humidified 37℃ incubator with 5% CO2. The cells were then characterized as previously described [1213]. Passages five to eight were used for experimentation.
2. Transfection of small interfering RNA into cells
The fibroblasts were serum-starved for 24 hours and transfected with 100 pmol small interfering RNA (siRNA) oligonucleotides targeted specifically to HDAC7 (Santa Cruz Biotechnology, Santa Cruz, CA, USA) by using Lipofectamine 2000 (GIBCO). In parallel, 100 pmol scramble siRNA was used as a control. After transfection, cells were plated and cultured for 48 hours in DMEM. The fibroblasts were then treated with 10 ng/mL TGF-β1 (R&D Systems Inc., Minneapolis, MN, USA) for 24 hours to detect the protein expression of plasminogen activator inhibitor-1 (PAI-1), fibronectin, collagen subtypes, smooth muscle α-actin, and HDAC7.
3. Reverse-transcriptase polymerase chain reaction
Total RNA was extracted from cultured cells with Trizol (Invitrogen, Carlsbad, CA, USA) according to the manufacturer's protocols. RNA was reverse-transcribed by use of the Reverse Transcription System (Promega, Madison, WI, USA) according to the manufacturer's instructions. Reverse-transcriptase polymerase chain reaction (RT-PCR) fragments were amplified by using the AccuPower PCR premix (Bioneer, Alameda, CA, USA). The PCR reaction was performed with denaturation at 94℃ for 1 minute, annealing at 60℃ for 30 seconds, and extension at 72℃ for 30 seconds (25 cycles). For the analysis of PCR products, 10 µL of each PCR reaction was electrophoresed on a 1% agarose gel and DNA bands were visualized with a ultraviolet illuminator (Image Station IS4000R system; Kodak, Rochester, NY, USA). Glyceraldehyde 3-phosphate dehydrogenase was used as an internal control. The primer sequences are listed in Table 1.
4. Western blot
Equal amounts of protein from whole-cell extracts (50 µg/lane) were electrophoresed on 12% sodium dodecylsulfate-polyacrylamide gels, transferred to nitrocellulose membranes, and probed with antibody against HDAC7 (1:100; Santa Cruz Biotechnology), PAI-1 (1:300; Abcam, Cambridge, UK), fibronectin (1:300; Abcam), collagen I (1:300; Abcam), collagen IV (1:300; Abcam), smooth muscle α-actin (1:300; Sigma-Aldrich), or β-actin (1:6,000; Abcam).
5. Fluorescent immunocytochemistry
The fibroblasts were cultured on sterile cover glasses (Marienfeld Laboratory, Lauda-Königshofen, Germany) and grown until nearly confluent. The cells were washed three times with PBS and then fixed in 4% paraformaldehyde for 10 minutes at 4℃ and in 100% methanol for 10 minutes at 4℃. Individual chambers were incubated with antibody to PAI-1 (1:300; Abcam), fibronectin (1:300; Abcam), collagen I (1:300; Abcam), collagen IV (1:300; Abcam), smooth muscle α-actin (1:300; Sigma-Aldrich), F-actin (1:300; Sigma-Aldrich), or Smad2/3 (1:200; Cell Signaling, Beverly, MA, USA) overnight at 4℃ in a moist chamber. After several washes with PBS, the chambers were incubated with fluorescein isothiocyanate-conjugated (1:300; Zymed Laboratories, South San Francisco, CA, USA) or tetramethyl rhodamine isothiocyanate-conjugated (1:300; Jackson ImmunoResearch Laboratories Inc., West Grove, PA, USA) secondary antibodies for 3 hours at room temperature. Mounting medium containing 4,6-diamidino-2-phenylindole (Vector Laboratories Inc., Burlingame, CA, USA) was applied to the chamber and nuclei were labeled. Signals were visualized, and digital images were obtained with a confocal microscope (FV1000; Olympus, Tokyo, Japan) under identical exposure settings.
RESULTS
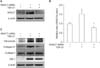
1. Differential gene expression of histone deacetylases in fibroblasts isolated from human Peyronie's disease plaque or from normal tunica albuginea
To examine the changes in gene expression of the HDAC isoforms, we performed RT-PCR. The mRNA expression of HDAC2, 3, 4, 5, 7, 8, 10, and 11 was higher in fibroblasts isolated from PD plaque than in fibroblasts isolated from normal TA. No detectable differences were noted in the gene expression of HDAC1, 6, and 9 (Fig. 1).
2. Histone deacetylase 7 knockdown inhibits extracellular matrix production induced by transforming growth factor-β1 in fibroblasts derived from human Peyronie's disease plaque
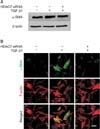
To determine the anti-fibrotic role of HDAC7, the siRNA approach was used. PD fibroblasts were transfected with siRNA specifically targeting HDAC7. Western blot analysis revealed that the treatment of PD fibroblasts with TGF-β1 induced HDAC7 expression and these expression was profoundly inhibited after treatment with HDAC7 siRNA (Fig. 2A, 2B).
3. Histone deacetylase 7 knockdown inhibits myofibroblastic differentiation induced by transforming growth factor-β1 in fibroblasts derived from human Peyronie's disease plaque
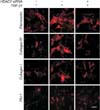
The expression of smooth muscle α-actin, a marker for myofibroblasts, at the protein level was determined with Western blot analysis. The treatment of PD fibroblasts with TGF-β1 resulted in an increase in smooth muscle α-actin expression, which was attenuated after treatment with HDAC7 siRNA (Fig. 4A). Fluorescent immunocytochemistry also revealed that HDAC7 siRNA inhibited TGF-β1-stimulated α-actin fiber formation (Fig. 4B).
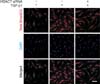
4. Histone deacetylase 7 knockdown inhibits nuclear translocation of Smad2/3 induced by transforming growth factor-β1 in fibroblasts derived from human Peyronie's disease plaque
TGF-β1 has been shown to induce the translocation of Smad2/3 proteins from the cytoplasm to the nucleus [14]. In order to evaluate whether HDAC7 inhibition affects on TGF-β1-induced nuclear shuttling of Smad2/3, we performed immunofluorescent staining of fibroblasts with antibody against total Smad2/3. The treatment of PD fibroblasts with TGF-β1 profoundly induced nuclear translocation of Smad2/3. HDAC7 siRNA reduced TGF-β1-induced nuclear accumulation of Smad proteins (Fig. 5).
DISCUSSION
Among the 11 HDACs tested in the present study, the expression of HDAC2, 3, 4, 5, 7, 8, 10, and 11 transcripts was higher in fibroblasts isolated from PD plaque than in fibroblasts isolated from normal TA. However, this result is limited by the small sample size, and further studies in a larger study population are required for further validation. It was also reported that the expression of both class I HDACs (HDAC1, HDAC2, HDAC3, HDAC8) and class II HDACs (HDAC4, HDAC 5, HDAC 7, HDAC 9) were significantly elevated in lung tissues from patients with idiopathic pulmonary fibrosis compared with those from non-diseased controls [15]. Moreover, siRNA mediated silencing of HDAC7 corrects the ΔF508 mutation in cystic fibrosis transmembrane conductance regulator that is responsible for premature lung failure and reduced lifespan in patients with cystic fibrosis [16]. HDAC7 is also known to be involved in the hepatic fibrosis by binding promoter region of hepatocyte growth factor (HGF) and limits the antifibrotic function of HGF [17]. These findings led us to investigate whether and how HDAC7 exerts its antifibrotic effects in fibroblasts isolated from human PD plaque.
Here, it was shown that siRNA-mediated knockdown of HDAC7 successfully ameliorated the TGF-β1-induced accumulation of extracellular matrix in human PD fibroblasts by blocking nuclear translocation of Smad2 and Smad3, the crucial step for TGF-β-mediated fibrosis, and by inhibiting TGF-β1-induced myofibroblastic differentiation.
In the present study, treatment of PD fibroblasts with TGF-β1 significantly induced HDAC7 protein expression. The specific gene knockdown of HDAC7 with siRNA significantly decreased the TGF-β1-induced accumulation of extracellular matrix proteins, such as fibronectin, PAI-1, collagen I, and collagen IV. Similar to the results from ours, silencing of HDAC7 in skin fibroblasts from patients with systemic sclerosis also decreased constitutive and cytokine (TGFβ1)-induced production of type I and type III collagen on both the mRNA and protein levels [11], suggesting HDAC7 as a potential therapeutic target in a variety of fibrotic diseases.
Accumulating evidences suggest that HDACs are involved in the cytokine-induced differentiation of fibroblast into myofibroblast [1318192021]. Treatment of rat renal interstitial fibroblasts with TSA, a nonspecific HDAC inhibitor, decreased the expression of smooth muscle α-actin and deposition of extracellular matrix [20]. We recently reported in human PD fibroblasts that inhibition of HDAC2 abrogated TGF-β1-induced transdifferentiation of fibroblasts into myofibroblasts [13]. Knockdown of HDAC4 is also known to inhibit TGF-β1-stimulated smooth muscle α-actin expression in normal human lung fibroblasts [18]. However, the role of HDAC7 in myofibroblastic differentiation is largely unknown. In the present study, treatment of PD fibroblasts with HDAC7 siRNA significantly reduced TGF-β1-induced fibroblast-to-myofibroblast transition. Because activation of fibroblasts into myofibroblasts is responsible for increased production of extracellular matrix, HDAC7 siRNA-mediated decrease in myofibroblastic differentiation is a key mechanism for halting fibrotic processes in the TA.
TGF-β1 is one of the most studied cytokine associated with PD and the expression of TGF-β1 and its downstream signaling pathway, such as Smad2 and Smad3 transcriptional factors, are known to be up-regulated in human PD plaque [122223]. TSA is known to inhibit nuclear translocation and DNA binding of Smad transcription factors in skin fibroblasts from patients with systemic sclerosis [10]. Similar to recent studies by us that showed a decrease in the nuclear translocation of Smad2 and Smad3 in human PD fibroblasts in vitro [13] and in PD rats in vivo [24], when HDAC2 was knockdown, treatment of PD fibroblasts with HDAC7 siRNA also prevented nuclear accumulation of Smad proteins. Therefore, suppression of the activation of Smad2 and Smad3 proteins is another mechanism by which silencing HDAC7 ameliorates fibrotic responses in PD fibroblasts. Further studies are required to determine the anti-fibrotic role of HDAC7 knockdown in PD models in vivo. It is also necessary to examine the efficacy of other HDAC isoforms in PD models.
In spite of distinctive physiologic function of each HDAC isoforms, most known HDAC inhibitors target multiple isoforms, which greatly limit their therapeutic utility. Therefore, dichotomizing individual function of HDAC isoforms by use of RNA interference technology may open new avenues for developing specific and safe treatment modality for PD.
CONCLUSIONS
The specific gene knockdown of HDAC7 in PD fibroblasts successfully attenuated TGF-β1-induced extracellular matrix production by inhibiting transdifferentiation of fibroblasts into myofibroblasts and by blocking activation of Smad2/3 pathway. Inhibition of HDAC7 with RNA interference may represent a promising epigenetic therapy for PD.


 XML Download
XML Download