

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Nutritional support or restrictions have been important issues for cancer control in recent decades. Milk is a very important food, and there have been many confusing and parochial reports on the relationship between milk intake and diverse cancers. Most epidemiologic studies [1,2,3,4,5,6], but not all [7,8], have reported an increase in prostate cancer risk with an increased milk intake. To elucidate the effect of milk on prostate cancer, numerous experimental studies have tried to identify hazardous ingredients in milk, such as calcium [6,9], estrogen [10], and insulin-like growth factor-1 (IGF-1) [11]. However, among the components of milk, protein has been enigmatic. A favorable effect of milk protein was the inhibition of mutation [12,13,14,15]. Several cell culture studies showed that milk protein could contribute to cancer prevention. The opposite results have appeared in recent epidemiologic studies [16,17]. The growth of prostate cancer cells was stimulated by milk protein [16,17]. Owing to these conflicting results, the potential role of milk protein in prostate cancer is still controversial.
Milk protein consists of 80% casein and 20% whey [18]. Casein has four subtypes: αs1-, αs2-, β-, and κ-casein [19]. α-Casein, a mix of αs1- and αs2-casein, is the main fraction of milk protein. Casein has potent antimutagenic effects proven through mutagen models [12,13,14]. Several animal experiments also showed an inhibitory effect of casein against mutagens [20,21]. Mice fed 20% casein diets had a significantly lower 1,2-dimethylhydrazine-induced colon cancer incidence than the control group [21]. The literature states that casein might prevent colon and breast cancer [20,21,22]. To date, however, the effect of casein itself on prostate cancer cells has never been investigated.
To establish the relationship between casein and prostate cancer, we evaluated the proliferation of immortalized prostate cells and diverse cancer cells including prostate cancer cells after treatment with casein and α-casein. Prior to a comparison between the casein-treated group and the control group, appropriate experimental conditions were established in prostate cancer cells PC3 and LNCaP.
MATERIALS AND METHODS
1. Cell lines and cultures
Human prostate cancer cell lines, androgen-independent PC-3 (ATCC, Manassas, VA, USA) and androgen-dependent LNCaP cells (ATCC, Manassas), were maintained in a Roswell Park Memorial Institute (RPMI) 1640 medium (Invitrogen, Carlsbad, CA, USA) supplemented with 10% fetal bovine serum (FBS), penicillin (100 U/mL), and streptomycin (100 mg/mL). Human A549 (ATCC, Manassas) lung cancer cells, SNU-484 (KCLB, Seoul, Korea) stomach cancer cells, and HEK293 (ATCC, Rockville, MD, USA) human embryonic kidney cells were maintained in the same manner. Human breast cancer cells (MCF7; ATCC, Rockville) were maintained in Dulbecco's modified Eagle's minimal essential medium (Invitrogen) with the aforementioned supplements. Immortalized RWPE-1 (ATCC, Manassas) normal human prostate cells were grown in bovine pituitary extracts (50 µg/mL), keratinocyte, and epidermal growth factor (5 ng/mL) under the same incubation conditions. The cells were cultured at 37℃ in a humidified atmosphere under 5% CO2 in air. The cells were plated first in 10% FBS; the growth medium was removed after 24 hours and replaced with a serum-free medium supplemented with NaOH, α-casein, and casein from bovine milk at concentrations of 0.1 or 1 mg/mL for 72 hours. The cells were diluted in an appropriate medium before each experiment.
2. Casein and experimental conditions
α-Casein and whole casein from bovine milk were purchased from Sigma-Aldrich (St. Louis, MO, USA). Each cell line (PC-3, LNCaP, MCF7, SNU484, A549, RWPE-1, or HEK293) was seeded in 12-well plates at a density of 1×105 cells/well under serum-free conditions. Each cell was treated with NaOH, α-casein, or casein from bovine milk at concentrations of 0.1 or 1 mg/mL on the first day only. After 3 days, proliferations of each cell line growth were measured by using an 3-(4,5-Dimethyl-thiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT) assay. Prior to the establishment of the aforementioned experimental conditions, each cell line was cultured under various conditions. The number of treatments (first day only vs. everyday), duration of experiments (2~7 days), conditions with or without serum supplement, and concentration of casein (α-casein) were tested. Because serum is also a nutrient like casein, we chose a serum-free condition as the final experimental condition. To ascertain the results, experiments were repeated at least three times.
3. Measurement of cell proliferation and morphological change
After treatment, cell viability was assessed by incubating cells with 0.5 mg/mL of MTT for another 4 hours. Formazan produced by viable cells was prepared in dimethyl sulfoxide. Colorimetric analysis was performed at 570 nm in a Multiskan enzyme-linked immunosorbent assay (ELISA) reader (Thermo, Vantaa, Finland). Cell viability was presented as a relative percentage of controls. In addition, cells were photographed using light microscopy.
4. Statistical analysis
A two-way analysis of variance (ANOVA) test was employed to compare the experimental groups with the control group, while results before and after treatments were compared using Tukey's comparison test. Statistical significance was determined at p<0.05. All statistical calculations were computed using PASW Statistics ver. 18 (IBM Co., Armonk, NY, USA).
RESULTS
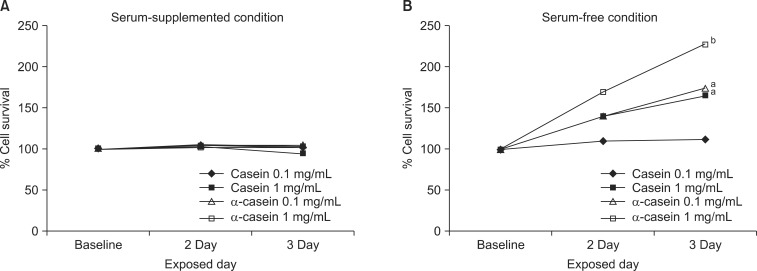
The changes in proliferation were evaluated after treatments with casein (or α-casein) for 2 to 3 days. There was no significant change in the proliferation of PC3 cells with FBS supplementation (Fig. 1A). The results in other cells were similar to the PC3 cell growth. Next, as FBS sometimes provides an artificial milieu, each cell was cultured in a serum-free medium and the changes in cell proliferation were investigated. Interestingly, casein induced a marked dose-dependent enhancement of cell proliferation in the PC3 and LNCaP cells (Fig. 1B). With the supplementation of 1 mg/mL of α-casein, the changes in the growths of PC-3 and LNCaP cells were 228% (p<0.001) and 134% (p<0.05), respectively. Similarly, the addition of 1 mg/mL of casein enhanced the growth to 166% (p<0.05) and 142% (p<0.05) proliferation in the PC-3 and LNCaP cells, respectively. However, there was no significant change in the growth of the other cells under the same experimental conditions.
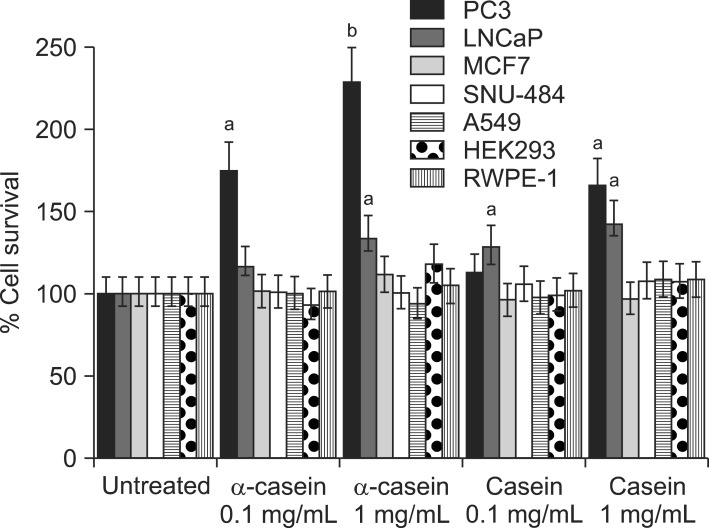
Fig. 2 shows the changes in the proliferation of each cell after 72 hours of serum-free culture. Treatment with α-casein induced marked dose-dependent increases in cell proliferation in the PC-3 and LNCaP cells. However, there was no change in the proliferation of the other cancer cells or immortalized prostate cells.
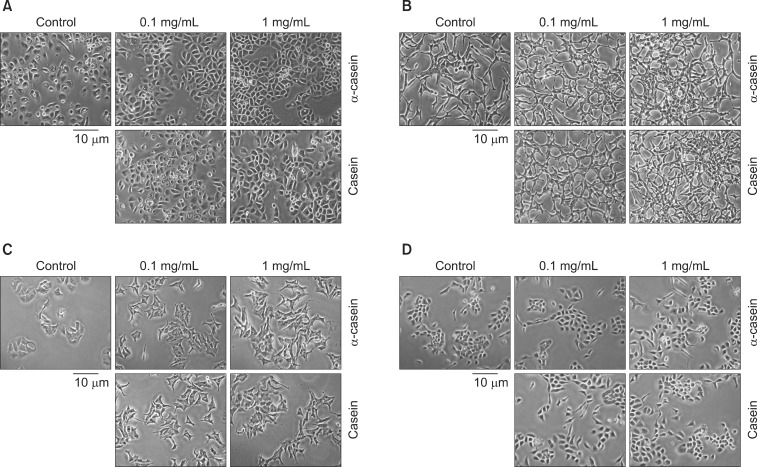
Fig. 3 shows representative morphological changes in each cell after exposure to α-casein and casein (0.1 and 1 mg/mL) for 72 hours. Under control conditions (NaOH alone), PC-3 cells appeared to have a typical phenotype, with round nuclei and homogeneous cytoplasm. PC-3 cells treated with casein or α-casein showed distinct morphological changes, increases in cell volume, cellular adhesion, and cell numbers. Likewise, LNCaP cells treated with casein or α-casein showed increased cellular adhesion. MCF7, SNU-484, and A549 cells treated with casein or α-casein showed no morphological changes compared with the untreated cells. RWPE-1 and HEK293 cells treated with casein or α-casein also showed an increase in cellular adhesion without any changes in cell volume or numbers.
DISCUSSION
Most epidemiologic studies have reported an unfavorable effect of milk on prostate cancer risk [1,2,3,5,8,23]. However, experimental results are diverse and complicated by several factors. First, the effects of the numerous components of milk remain unproven. Second, most components of milk interact with other components including hormones, growth factors, and minerals. Third, the results obtained in vitro should be replicable in animals or an in vivo study. Fourth, to understand the effect of dietary compounds on specific cells in a living body, digestion and their metabolites must also be considered.
Recent studies revealed that milk protein plays a key role in the development and modulation of prostate cancer cell proliferation [16,17,23]. Tate et al [16] reported that cow's milk stimulated the growth of prostate cancer cells (LNCaP) almost as much as digested whole milk. Like our results, neither casein nor digested milk increased the growth of breast cancer cells (MCF-7). Nielsen et al [17] evaluated the effect of whey protein, a second fraction of milk protein, on prostate cancer cell (PC-3) growth. While whey had an anti-proliferative effect on breast cancer cells (MCF-7), this protein had a stimulatory effect on prostate cancer cell (PC-3) growth under the control of the estrogen concentration. Our study also showed the proliferative effect of casein and α-casein on prostate cancer cells (PC-3). However, there is no significant change in the growth rate of breast cancer cells (MCF-7). Nevertheless, milk protein increases the proliferation of prostate cancer cells.
Milk includes multiple nutritional factors, carbohydrates, lipids, amino acids, and minerals [24]. Naturally, most milk proteins including casein can promote the growth of both normal cells and cancer cells. However, this study showed no proliferation of immortalized normal human prostate cells (RWPE-1). In addition, the growth of other cancer cells, human lung cancer (A549) and stomach cancer cells (SNU484), were not stimulated. We can speculate on the basis of these results that casein provided cancer-specific proliferating factors, not simple nutritional support.
The difference in the growth rate between PC3 and LNCaP is an intriguing finding. The growth of the PC3 cells was promoted more than that of the LNCaP cells by casein (or α-casein). However, no definite reason could be identified due to a lack of previous work. Note that androgen-sensitive LNCaP cells and -independent PC3 cells have different genetics, apoptotic behaviors, cell viability, and signaling pathways [25,26].
Unlike that of prostate cancer, the growth of breast cancer cells was reported to be suppressed by casein in the literature. Bonuccelli et al [22] showed that α-casein could significantly inhibit the growth and metastasis of one murine mammary tumor cell line (Met-1 cells) and two human breast cancer cell lines (MCF10A-H-Ras and MDA-MB-231 cells). In addition, α-casein mediated its tumor suppressor effects through the activation of STAT1 signaling. They speculated that α-casein could provide a 'differentiation' therapy for breast cancer.
Several authors have evaluated the effects of various states of casein on colon cancer. However, these results were discordant. Corpet and Chatelin-Pirot [27] reported that cooked casein could promote colon cancer in rats, perhaps because of mucosal abrasion. However, in an in vitro study, casein hydrolysates had an inhibitory effect on the viability and growth of colon cancer cell lines [28]. Recently, casein phosphopeptides, a family of bioactive peptides derived from the digestion of casein, were shown to modulate proliferation and apoptosis in intestinal adenocarcinoma cell lines [29].
To date, the theory that circulating IGF-1 concentrations are positively associated with an increased prostate cancer risk in humans has been the commonly accepted. Kimura et al [30] found that 9% of the IGF-1 fed to mice survived digestion and could be recovered intact out of the bloodstream; this figure increased to 67% when IGF-1 was fed together with casein. These observations provide very strong circumstantial evidence of increased levels of serum IGF-1 through excessive milk intake. Consequently, we understand that dietary casein might boost IGF-1 levels or prostate cancer cell proliferation. For this reason, the present study compared the effects of casein on the proliferation rates of various cancer cells. Despite greater proliferation in prostate cancer cells, there was no significant difference in the level of IGF-1 (data not shown). The addition of IGF-1 did not result in a further enhancement of prostate cancer cell growth [16]. These results suggest that casein promotes the proliferation of prostate cancer cells without any effect on IGF-1.
Casein cannot be absorbed directly from the digestive system. However, casein and α-casein have been detected in various conditions and tissues, even serum. No obvious mechanism of how casein might be transported from the intestines to the body tissues or cancer cells has yet been identified. Although casein promoted the growth of cancer cells under serum-free conditions in this study, it is not clear whether dietary casein could have an effect on prostate cancer cells in vivo. Further experiments on the molecular mechanisms of casein induced proliferation in prostate cancer cells and in vivo studies should be conducted.
CONCLUSIONS
Major milk proteins, α-casein and total casein, promoted the proliferation of PC-3 and LNCaP prostate cancer cells under serum-free conditions, but did not elicit any changes in the proliferation of other cancer cells or immortalized normal prostate cells. Furthermore, casein and α-casein showed dose-dependent proliferating properties. These effects of casein were not associated with IGF-1. To understand the relationship between proliferating prostate cancer cells and casein, a study on the molecular mechanisms of casein induced proliferation in prostate cancer cells as well as in vivo studies should be conducted.
 XML Download
XML Download